INTRODUCTION
A stroke is a general term for a sudden, focal neurological deficit caused by an abnormality in blood flow to the brain [18]. An ischemic stroke occurs when the blood supply to a localized area of the brain is reduced, resulting in dysfunction in that area. The central nervous system affected by an ischemic stroke develops aphasia, vision defects, memory deficits, and impaired thinking. When the blood vessels to the brain are compromised, the supply of glucose, the brain’s energy source, is compromised [22]. When glucose is the only source of energy, brain tissue is more susceptible to necrosis. This damage to brain tissue caused by abnormalities in cerebral blood flow can lead to symptoms of cognitive impairment. Ischemic stroke triggers an ischemic cascade [5,6]. Ischemic conditions induce the production of oxygen free radicals and reactive oxygen species, which can damage cells [25].
Some patients who have suffered a stroke will have residual disability even after receiving treatment [15,23]. More than half of patients receiving acute therapy progress to long-term disability. With the development of acute therapies, survival rates for stroke patients have improved [8,14,29]; however, post-stroke cognitive impairment and dementia remain prevalent [11,13,19,26,27], with 70% of stroke survivors experiencing cognitive impairment among their sequelae [28].
In ischemic stroke, there are several well-studied biomarkers that can identify neuroinflammation from the ischemic cascade, including interleukin (IL)-1β, IL-6, and tumor necrosis factor [3]. On the other hand, biomarkers for neural damage after ischemic stroke are less well studied. The expression of inflammation-related factors is upregulated in ischemic stroke conditions. However, an increase in inflammatory factors does not necessarily mean that an ischemic stroke will occur. The authors were convinced of the need for a neural damage biomarker that could demonstrate neural damage along with the progression of the ischemic cascade. A study was conducted to find biomarkers of neural damage after ischemic stroke.
MATERIALS AND METHODS
Experimental animal and research design
All experiments were conducted under the approval of IACUC (SCH22-0097). The housing area of the mice used in the experiments was subjected to a 12-hour light/dark cycle (7:00 a.m. to 7:00 p.m.), the room temperature was maintained at 23°C±1°C, and the room humidity was maintained at 50%±5%.
Experimental animal and classification criteria
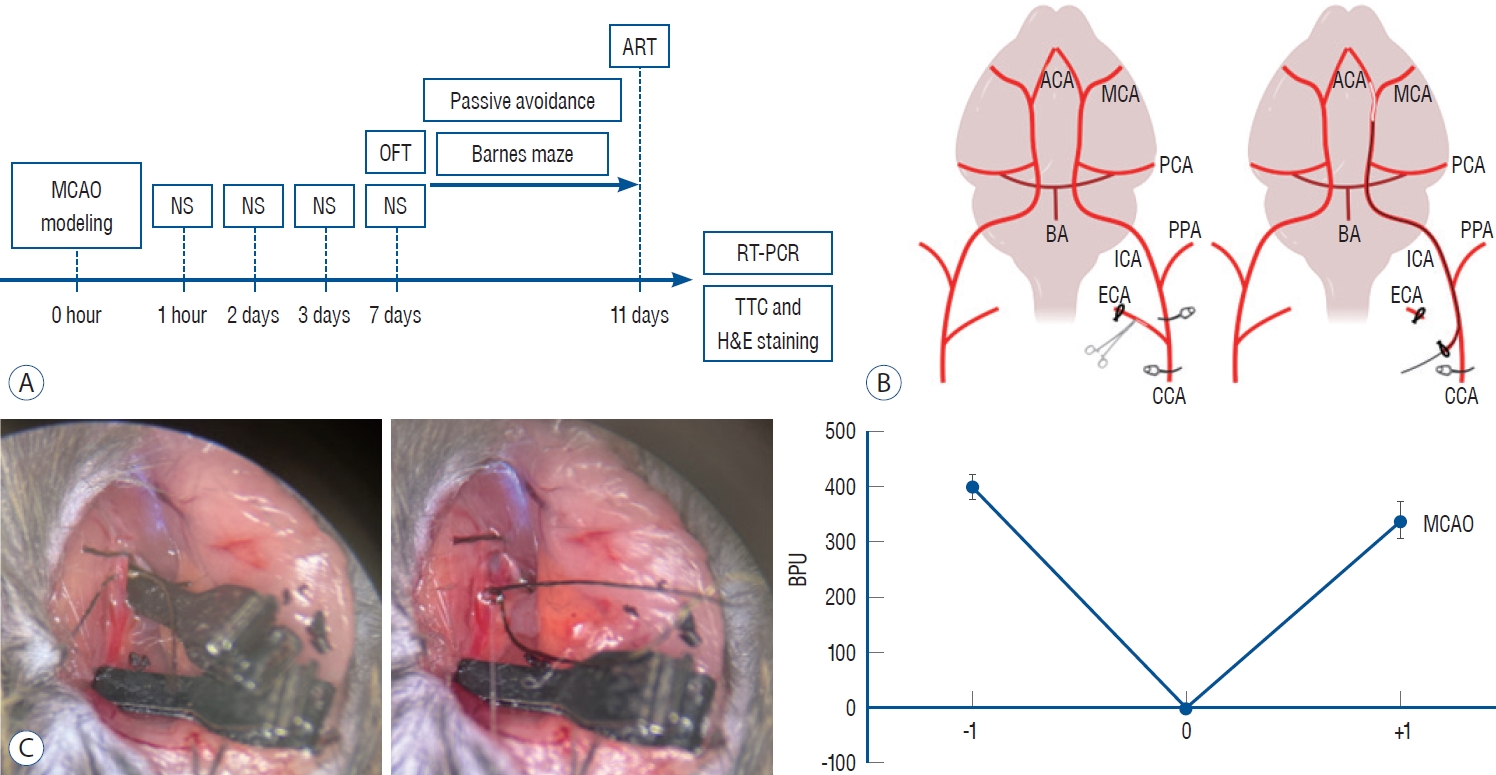
The experimental animals used in the study were house mice (Mus musculus) supplied by RaonBio (Yongin, Korea). All mice were male, C57BL/6J strain, weighing 25 g and aged 8 weeks, and were randomly assigned to groups. The groups were designed as a healthy wild-type (WT, n=6) control group, a lowseverity middle cerebral artery occlusion (MCAO) 1 (n=6) group, and a high-severity MCAO2 (n=6) group based on neurological scoring after ischemic stroke. The classification criteria were based on previous studies that identified the most pronounced neurological impairment at 48 hours after MCAO, so individuals with a 48-hour neurological score of less than 4 were classified as MCAO1, the low-impairment group, and individuals with a score of more than 4 were classified as MCAO2, the high-impairment group.
MCAO surgery
The experimental group performed Longa et al. [20]’s MCAO method to implement a mouse model of sequelae after ischemic stroke. Blood flow was measured by noninvasive laser Doppler (OxyFlo™ Pro; OXFORD OPTRONIX, Banbury, UK) in three phases : before, during, and after, to determine normal blood flow, blood flow during MCAO, and blood flow after achieving reperfusion.
After induction of anesthesia at 2.5%, isoflurane was maintained at 1.5%. The body temperature of the mouse was maintained at 37°C degrees with a heating pad. Mouse MCAO was performed to induce reperfusion after ischemic stroke. After vascular clamping the left common carotid artery (CCA), left internal carotid artery (ICA), and left external carotid artery (ECA) of the mouse, the MCAO filament was inserted through the incision after temporarily ligating the left ECA. Place the MCAO filament past the ICA and into the M1 region of the MCA to achieve occlusion. After 30 minutes of MCAO, the filament was removed to achieve reperfusion, and the incision was sutured. After surgery, the mice were provided with a heat irradiator and a recovery diet.
Post-operative care and neurological score
After waking up from anesthesia, the mice were given a recovery diet. Neurological scores were then measured to determine the extent of neural damage of post-ischemic stroke and mouse health. All mice were measured, and two measures were performeD : torso turning and forepaw grip strength. Measurements were taken immediately after awakening from anesthesia and on days 1, 2, 3, and 7 after MCAO. The torso rotation test measures the degree to which the mouse’s body curves in the air in the direction opposite to the side on which the hemiparesis is induced by grabbing the mouse’s tail and lifting it. A score of 3 is given for an immeasurable amount of body flexion, 2 for severe body flexion, 1 for mild body flexion, and 0 for no movement at all. For the forefoot strike test, the athlete is asked to hold the bar with both forefoots, and the point at which the forefoot that induces unilateral paralysis falls before the contralateral forefoot. A score of 3 is given for unmeasurable hemiparesis, 2 for a very fast fall of the hemiparetic forefoot before the contralateral forefoot, 1 for a slow fall of the forefoot with mild hemiparesis, and 0 if both forefoots have no problem gripping the bar. Each score for torso turning and grip strength was added together to give everyone a rating from 0 to a maximum of 6.
Behavioral tests
All groups were subjected to behavioral tests to analyze behavioral changes in activity and cognitive memory due to ischemic stroke. Behavioral analysis began on day 7 after MCAO and lasted for a total of 5 days (day 11 after MCAO), including an adaptation period, in which experiments were designed from low to high external stimuli to account for the continuous stress of the mice. Each behavioral experiment was video recorded for analysis.
Open field test
A cube-shaped transparent acrylic box (x×y×z, 45×45×45 cm) with an open top is divided into a central zone and a peripheral zone corresponding to 50% of the area of the horizontal axis (x) and vertical axis (y), and the mouse is introduced from the central zone and allowed to move freely for 10 minutes. After the end of the experiment, the total distance traveled is analyzed using behavioral analysis software.
Barnes maze test
Designate 18 localized zones in a circular field 92 cm in diameter, with only one of them containing the mouse’s usual diet. Divide the field into quadrants, allocating 25% of the field to each. The mice start in the center of the field and have three minutes to explore freely, ending early if they reach the food source before the 3 minutes are up. After 3 days of adaptation, analyze latency and distance to target on the fourth test day.
Adhesive removal test
Attach a single-sided tape (x×y, 3×2 mm) to one forepaw of a mouse and place it in a translucent cage with no bedding to allow it to behave freely. Observe the mouse for up to 2 minutes and analyze the time it takes for the mouse to remove the tape. Measure both forepaws, starting with one side and switching to the other side when finished.
Analyzing camera tracking
During the behavioral experiments, the recorded videos were analyzed with Smart 3.0 Video Tracking System software (Panlab, Barcelona, Spain). Smart 3.0 detects movement by comparing the current video with a reference image. Behavioral differences between each mouse individual were analyzed and quantified by precise automatic measurements of experimental time, mouse position, distance traveled between zones, mouse occupancy time, and the number of times the mouse crossed between zones.
Real time-polymerase chain reaction (RT-PCR)
At the end of the behavioral tests experiment, the group was randomly divided into two groups : one for histological analysis and the other for messenger RNA (mRNA) level analysis.
Brain tissue harvesting
Mice were euthanized by cervical dislocation after induction of isoflurane respiratory anesthesia. After confirmation of euthanasia, the neck was decapitated with surgical scissors to obtain the head separately, the scalp was rolled up to the front of the head to obtain the skull, forceps were inserted subdural and gently lifted outward to remove bone, and whole brain tissue was obtained. The brain tissue was transferred to a microtube, and 1 mL of easy-BLUE (trizol) was added. Immediately, the brain tissue and easy-BLUE were mixed evenly using a homogenizer.
RNA extraction
Add 200 µL of chloroform to the homogeneously mixed tissue and solution, mix, and run the centrifuge at 13000 rpm, 4°C for 10 minutes. After 10 minutes, when the protein, DNA and RNA layers are separated, take 400 µL from the RNA layer, which is the clear supernatant, transfer it to a 1.5 mL microtube and add equal volume of isopropanol (1 : 1) to precipitate the RNA for 10 minutes. After precipitation, centrifuge again at 13000 rpm, 4°C for 5 minutes. Check the RNA pellet after centrifugation and carefully remove only the supernatant. After removal, add 1 mL of ethanol diluted in 75% nuclease free water (NFW) to wash away the remaining solvent. Centrifuge again at 10000 rpm, 4°C for 5 minutes, remove the supernatant, and dry the pellet until it is dry. Add 20 µL of pre-heated NFW and quantify the amount of RNA via nanodrop at 1000 ng/mL, calibrating all individuals equally.
cDNA synthesis
Add the calibrated RNA dilution to the RNA dilution, NFW, and 2 µL of master mix according to the standard protocol provided by All-In-One 5X cDNA Master Mix (CellScript, Madison, WI, USA) to a final volume of 10 µL. After addition, proceed with cDNA synthesis by reverse transcription according to the standard protocol for cDNA synthesis provided by the T100 Thermal Cycler (BIO-RAD, Contra Costa, CA, USA). Upon completion of synthesis, add 90 µL of NFW to dilute to 1/10.
RT-PCR
In one well of a 96-well plate, dispense 14 µL of primers, SYBR green, and NFW corresponding to the transcripts you want to target according to the standard protocol, then add 1 µL of cDNA for everyone for a total volume of 15 µL. Place a PCR sealing film on top of the 96 wells to prevent the contents of each well from evaporating and foreign substances from entering. Load the plate into the CFX96 Real-Time PCR System (BIO-RAD) and proceed with the polymerase reaction according to the PCR protocol. The PCR protocol is 95°C for 15 minutes, followed by 40 cycles of 95°C for 10 seconds, 60°C for 15 seconds, and 72°C for 20 seconds.
Triphenyltetrazolium chloride (TTC) staining
After euthanizing the mice, follow the same procedure for harvesting brain tissue for RNA extraction before homogenization. Obtain complete brain tissue, embed it in brain matrix, shape it, and cut it with a razor blade to obtain brain tissue sections at 2 mm intervals. The obtained brain tissue sections are completely immersed in a 2% dilution of TTC, wrapped in foil, and incubated in an incubator at 37°C for 30 minutes. After incubation, the unstained area (white) of the total tissue area was measured as a percentage (%) using Image J software (National Institutes of Health, Bethesda, MD, USA).
Hematoxylin and Eosin (H&E) staining
Once the mouse is unconscious under respiratory anesthesia, make an incision between the ribs and abdomen to access the heart. Once visualized, insert the needle into the convex convergence at the tip of the heart to inject phosphate buffered saline (PBS) via syringe. Once the needle is inserted, slowly inject PBS to achieve perfusion. Once perfusion is achieved, the brain is obtained through the same incision and the tissue is fixed by complete immersion in 4% paraformaldehyde. The fixed brain tissue was sectioned to a thickness of approximately 2-3 mm and subjected to tissue processing (STP120 Spin tissue Processor; Especialidades Médicas MYR, S.L., Tarragona, Spain). The processed tissue was sectioned using a sectioning machine (Finesse ME Microtome; Thermo Fisher Scientific, Waltham, MA, USA) to a thickness of approximately 3-4 µm, and the sections were attached to slides for drying, deparaffinized in xylene for 3 steps/3 minutes, hydrated in alcohol for 4 steps/2 minutes, hematoxylin for 10 minutes, water for 3 minutes, eosin for 1 minute 40 seconds, alcohol for 4 steps/1 minute, cleared in xylene for 3 steps/3 minutes, and embedded to prepare H&E sections.
Statistical analysis
Quantitative data obtained through behavioral tests analysis, mRNA level analysis, TTC staining were graphed and statistically analyzed using GraphPad Prism 8 software (GraphPad Software, Boston, MA, USA). Statistically differences between each control and experimental group were tested for significance using one-way analysis of variance.
RESULTS
MCAO blocks cerebral blood flow, leading to ischemic stroke
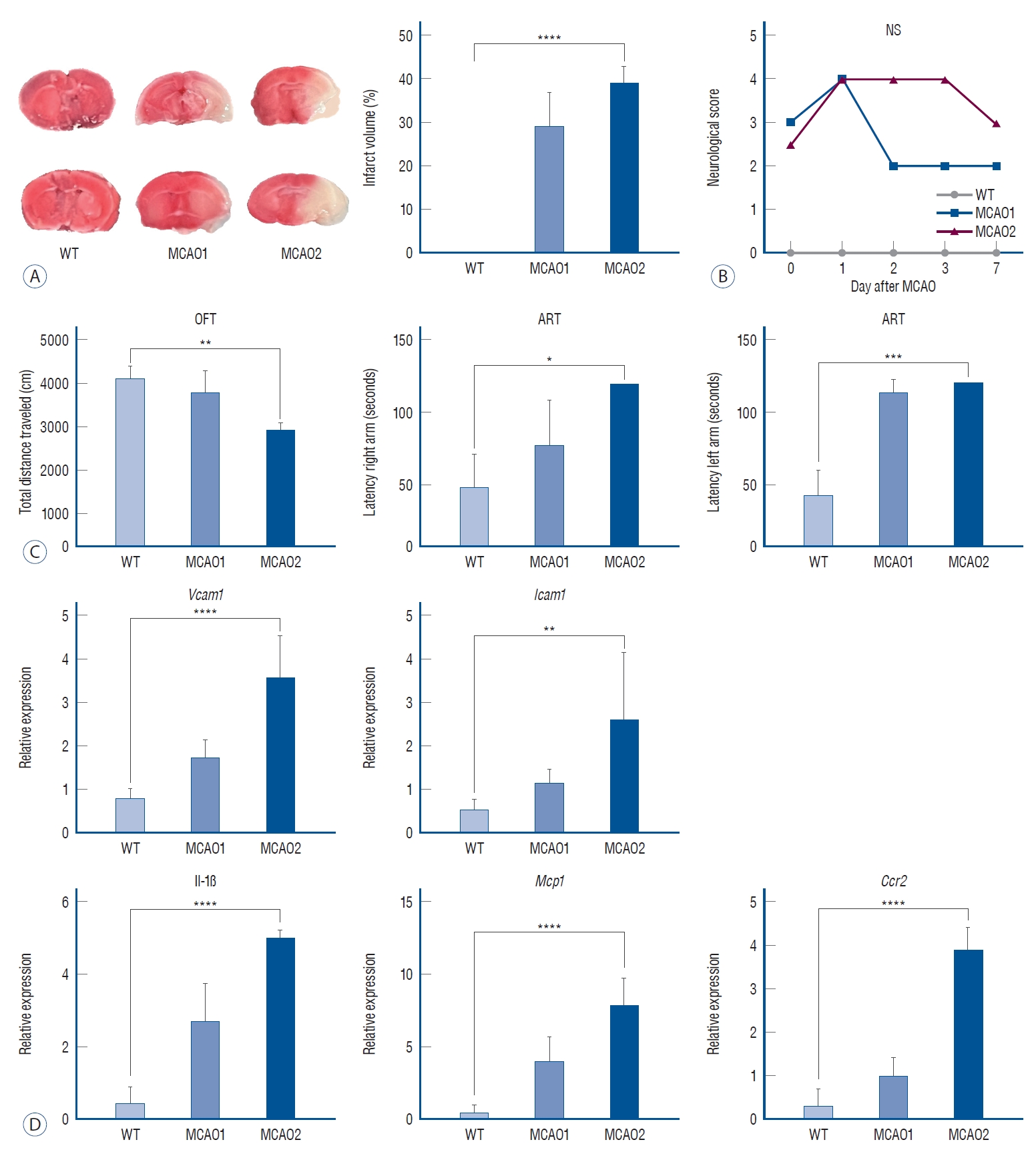
In the MCAO group, reperfusion was performed after Longa MCAO (Fig. 1). Immediately before occlusion of the M1 segment of the middle cerebral artery, blood flow was measured by noninvasive laser doppler, which revealed an average blood flow of 400 blood perfusion unit (BPU). During the MCAO, researcher found 0 BPU, and when researcher achieved filament retraction and blood reperfusion, researcher found blood flow reperfusion between 300 BPU and 400 BPU.
For all groups, brain tissue obtained after sacrifice at the end of the behavioral analysis was subjected to TTC staining for cerebral infarct localization, and cerebral infarction was confirmed in the MCAO1 and MCAO2 groups (Fig. 2A). No cerebral infarction was identified in WT (0%), 29.02%±7.74% in MCAO1 and 38.94%±3.74% in MCAO2, with MCAO2 having the highest infarct volume expression (p<0.0001). MCAO2 showed a higher infarct volume than MCAO1 (p=0.0065). MCAO2 scored higher on neurological scoring measures, confirming the damaged brain (Fig. 2B).
MCAO group expressed neuroinflammatory brain damage due to ischemic cascade
The MCAO group that experienced ischemic stroke had neuroinflammatory damage from the ischemic cascade (Fig. 2D).
RT-PCR performed to determine differences in mRNA level expression showed that the mRNA levels of vascular cell adhesion molecule 1 (Vcam1), which contributes to immune cell adhesion [9], were 0.81±0.22 in WT, 1.74±0.21 in MCAO1, and 3.61±0.95 in MCAO2 (p<0.0001). The mRNA level of Vcam1 in MCAO2 was higher than in MCAO1 (p=0.0013).
The mRNA level of intercellular adhesion molecule 1 (Icam1), which contributes to immune cell adhesion [9], showed expression levels of 0.54±0.24 in WT, 1.15±0.33 in MCAO1, and 2.63±1.53 in MCAO2 (p=0.0037). The mRNA level of Icam1 in MCAO2 was higher than in MCAO1 (p=0.0423).
The mRNA level of the pro-inflammatory cytokine [17] IL-1β was 0.44±0.44 in WT, 2.69±1.06 in MCAO1, and 5.02±0.20 in MCAO2 (p<0.0001). The mRNA level of IL-1β in MCAO2 was higher than in MCAO1 (p=0.0004).
The mRNA level of monocyte chemoattractant protein 1 (Mcp1), a chemokine receptor that recruits monocytes to the site of inflammation [4], showed expression levels of 0.48±0.50 in WT, 3.98±1.72 in MCAO1, and 7.86±1.85 in MCAO2 (p<0.0001). The mRNA level of Mcp1 in MCAO2 was higher than in MCAO1 (p=0.0038).
The mRNA level of C-C chemokine receptor 2 (Ccr2), which is involved in monocyte infiltration in inflammatory diseases, showed expression levels of 0.33±0.37 in WT, 1.02±0.39 in MCAO1, and 3.89±0.51 in MCAO2 (p<0.0001). The mRNA level of Ccr2 in MCAO2 was higher than in MCAO1 (p=0.0001).
Locomotor activity and sensorimotor changes in MCAO
The MCAO group exhibited symptoms of locomotor activity disorders (Fig. 2C). In terms of total distance traveled (cm) in the open field test, WT showed a total distance of 4102±621 cm, MCAO1 showed a total distance of 3771±490 cm, and MCAO2 showed a total distance of 2908±188 cm (p=0.0015). The total distance of MCAO2 was lower than MCAO1 (p=0.0024).
In the adhesive removal test for sensorimotor measures, WT showed a latency of 49±23 seconds, MCAO1 88±45 seconds, and MCAO2 120±0 seconds in the right forepaw latency region (latency right arm) (p=0.0168). Right forepaw adhesive removal latency was longer in MCAO2 than in WT (p=0.0055).
In the left forepaw latency measure (latency left arm), WT showed a latency of 43±17 seconds, MCAO1 114±9 seconds, and MCAO2 120±0 seconds (p=0.0004). There was a statistically different. Left forepaw adhesive removal latency was longer in MCAO2 than in WT (p=0.0011).
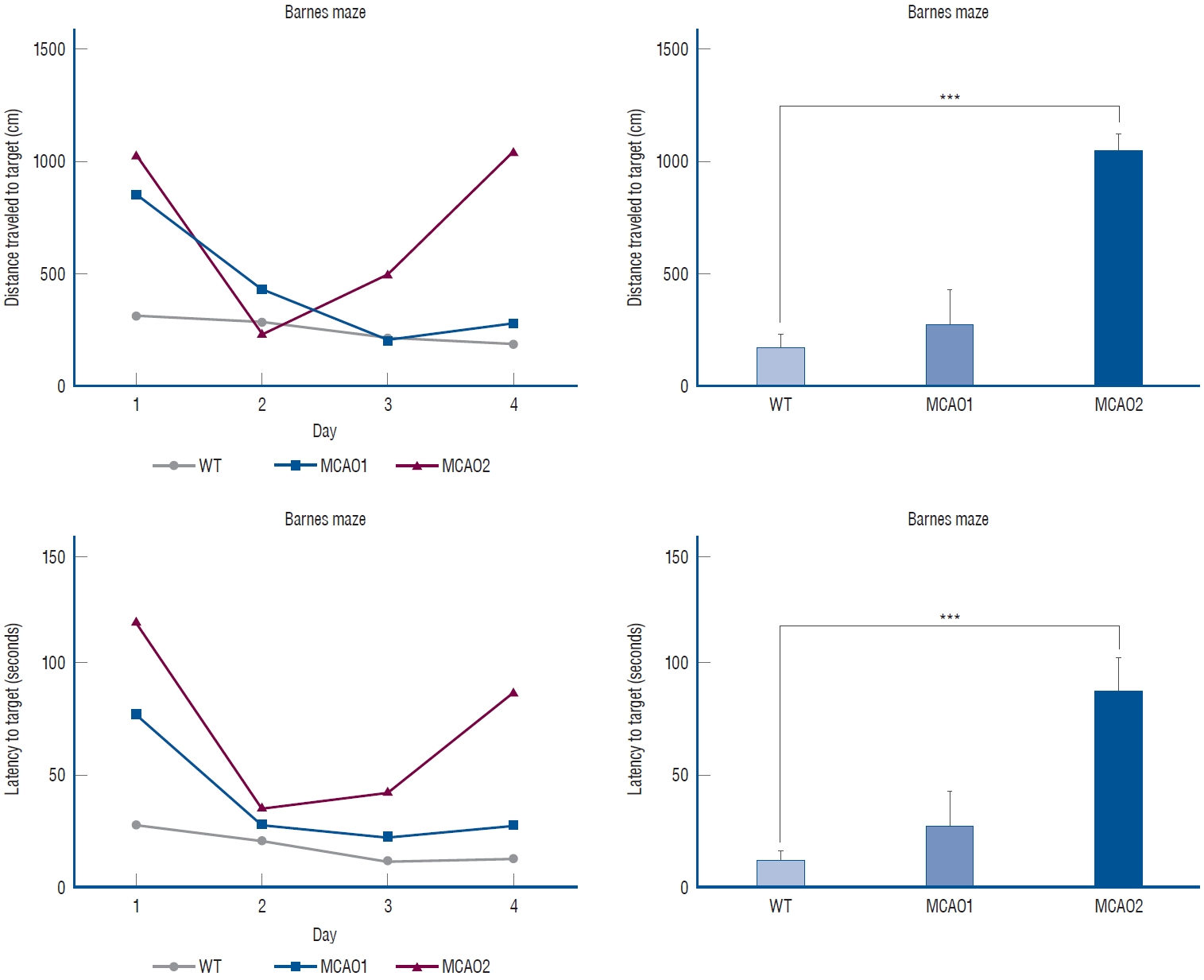
MCAO expressed impairment in positive long-term cognitive memory
The MCAO group exhibited long-term cognition memory impairment (Fig. 3). In the distance to target (cm), WT had 178±131 cm, MCAO1 had 276±216 cm, and MCAO2 had 1051±102 cm (p=0.0004). In distance to target, MCAO2 traveled a longer distance than MCAO1 (p=0.0442).
In the latency to target (seconds), WT showed a latency of 13.3±7.5 seconds, MCAO1 showed a latency of 27.9±21.1 seconds, and MCAO2 showed a latency of 87.9±21.2 seconds (p=0.0007). MCAO2 showed longer latency than WT in latency to target (p=0.0002).
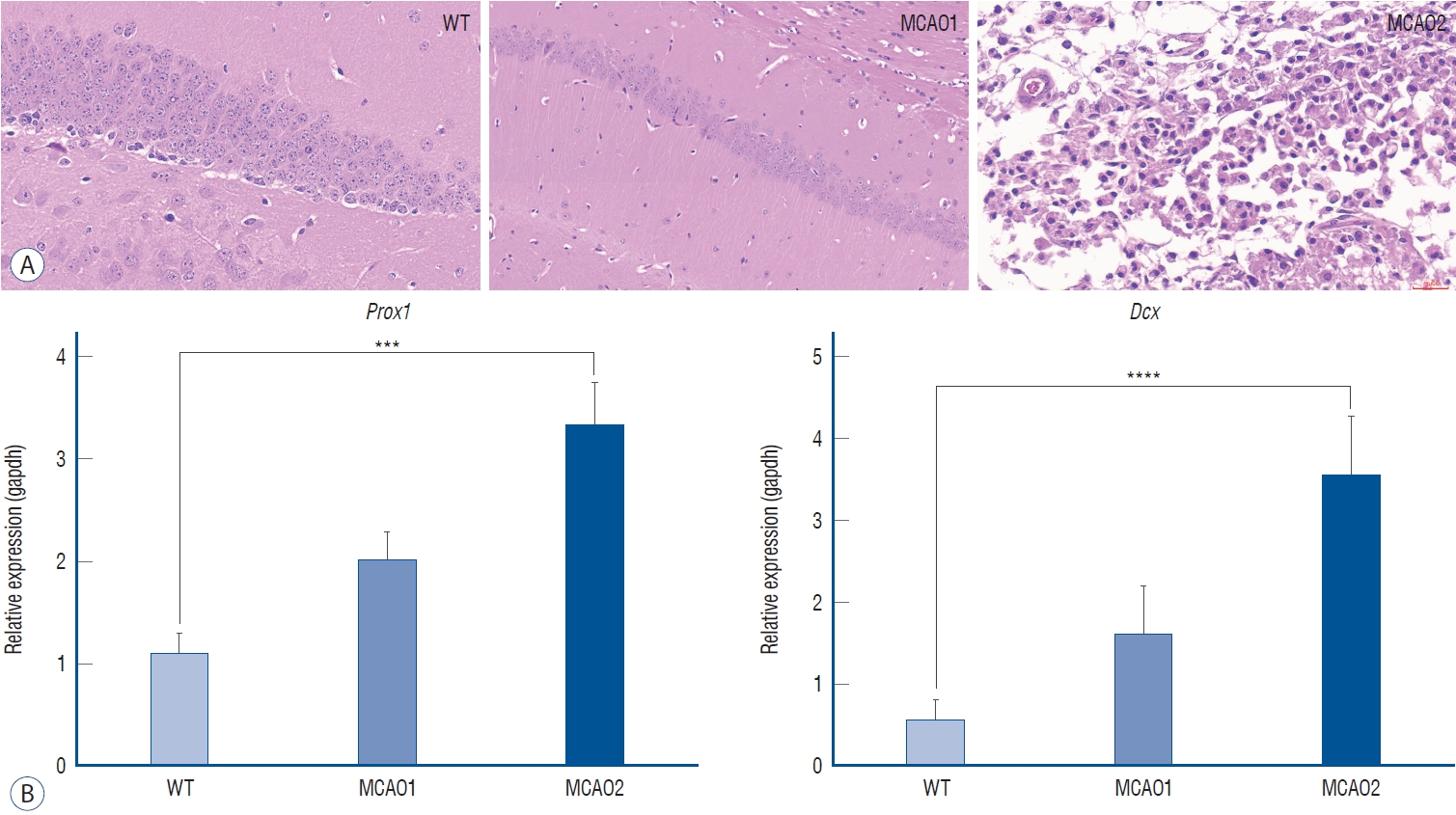
Altered mRNA levels of prospero homeobox 1 (Prox1) and Doublecortin (Dcx) and neural damage in MCAO
The MCAO group had confirmed neural cell damage (Fig. 4A). In healthy WT, neural cell arrangement was even, and no apoptosis was identified, MCAO1 had no apoptosis but hippocampal arrangement atrophy was identified, and MCAO2 had extensive apoptosis.
DISCUSSION
Increased mRNA level expression of Prox1 and Dcx may be a marker of neuronal damage (Fig. 5). Among the behavioral disorders after ischemic stroke, impairment of long-term cognitive memory is present. The formation of memories involves the Papez circuit and the limbic system [1,31]. Explicit memories are conscious, intentional memories that are generated by information and experiences [30]. The hippocampus, amygdala, striatum, and others are involved in motivation, emotion, learning, and memory. Damage to the hippocampus can affect spatial memory and cognition in general [7]. Prox1 is expressed in the early hippocampus [10]. The expression of Prox1 in the severe MCAO2 group was the highest compared to the MCAO1 group, and tissue H&E staining to identify neural damage confirmed brain tissue damage, with consistent results in the Barnes maze test. It is hoped that Prox1 and Dcx can be utilized as a novel predictor of neural damage. Prox1 is specifically expressed in the dentate gyrus [16], which is part of the hippocampal synaptic circuitry, and contributes to the formation of new memories and spontaneous exploration [10]. The authors found that Prox1 has higher mRNA expression in higher levels of neural damage, which is associated with worse long-term cognitive function. This study found that MCAO2 with higher severity had the highest mRNA expression levels of Prox1. Consistent with H&E staining confirming hippocampal apoptosis in the MCAO group, it predicts that the increased mRNA level expression of Prox1 is an early hippocampal marker and is expressed by hippocampal damage. Dcx is a protein expressed in neural progenitor cells and immature neurons. In healthy adult neurons, downregulation of Dcx is observed along with the expression of neuronal nuclear [2]. The Dcx is a neurogenesis marker, which is increased in cerebral ischemia [21]. When this research compared the extent of neural damage in high-severity MCAO2 and low-severity MCAO1 by tissue staining, it found lower expression of Prox1 and Dcx, consistent with lower neural damage in MCAO1.
Elevated mRNA expression levels of Prox1 and Dcx were associated with deterioration of locomotor activity and upregulation of neuroinflammation-related biomarkers. TTC staining was used to measure brain infarct volume in the MCAO groups, and the MCAO2 group, which had the highest severity of neural damage, had the highest brain infarct volume. The expression of symptoms of cerebral infarction and hemiplegic disorders appears to be caused by cerebral ischemic damage by the ischemic cascade after stroke, which is the same in human ischemic stroke. The lack of oxygen and energy supply due to ischemia leads to mitochondrial death and apoptosis. Vcam1 and Icam1, adhesion molecules that help monocytes migrate, were found to be upregulated in the MCAO2 group. Higher mRNA levels of IL-1β, Mcp1, and Ccr2, inflammationassociated chemokines and cytokines, were found in the MCAO2 group. Behavioral tests confirmed behavioral impairment in the MCAO group. In the open field test, the MCAO2 group showed less activity than the MCAO1 group. In the adhesive removal test, the MCAO2 group had more difficulty removing the tape from both forepaws and reached a delayed maximum time compared to the MCAO1 group. The differences were even more pronounced in the Barnes maze test, which checks for positive long-term cognitive memory. From the first day of preliminary trials, the MCAO2 group spent more distance traveled and latency to reach the destination than MCAO1, and on the day of the final analysis measurements, they experienced many errors, which increased the distance traveled and latency.
There are limitations to the study. While this research has identified mRNA level expression differences to identify ischemic cascades and neural damage, this research lack confirmation that there are differences in regional expression in the form of functioning proteins or cells. It would be an interesting follow-up study to apply ischemic stroke countermeasures, such as thrombolytic or vasodilator drugs, or surgical treatment to the animal models in this study to see if mRNA expression of Prox1 and Dcx is reduced, and cognitive function is restored.
CONCLUSION
The severity of behavioral disorders following ischemic stroke was correlated with the severity of histological damage in the brain and upregulation of biomarkers. Increased mRNA level expression of Prox1 and Dcx is proportional to the severity of neural damage after stroke. This study suggests Prox1 and Dcx as biomarkers of neural damage.