INTRODUCTION
Radiotherapy is a treatment modality for primary central nervous system (CNS) tumors and metastases [24]. While radiation therapy affects tumor cells, it also affects normal brain cells. The effects of cranial radiation on cognitive functions determine the quality of life. Especially in treating brain tumors, quality of life is as important or should be as important as living a long time. Radiation can cause impairment of functions such as thought, memory, concentration, and speech and reduce the quality of life [23].
The use of growth factors in treating brain injuries has recently become popular. Granulocyte colony stimulating factor (G-CSF) was first used in treating leukopenia and was later used in animal stroke models. In these brain models, the preventive effect of G-CSF on brain damage has been demonstrated, and positive outcomes have been achieved [31,33]. Moreover, previous studies showed that intraperitoneal or intravenous administration of G-CSF had positive effects in animal models [1,40]. Various studies have shown that single G-CSF treatment modalities can also be used in traumatic brain injury for cognitive function recovery [32] or cellular proliferation with motor function recovery [27].
Reactive oxygen species (ROS) are delicately regulated in brain tissue under physiological settings by the interaction of pro-oxidant and antioxidant mechanisms. Even Nevertheless, there is an increase in ROS generation in cases of brain damage [38]. When there is inflammation in the brain, cytokines produced by neurons and glial cells control the inflammatory process, and nuclear factor kappa-B (NF-κB) is involved in processes that affect neuronal function, including the immune response, cell cycle, survival, proliferation, and development, plasticity, and memory formation.
Animal models are essential to evaluate brain functions, such as anxiety response, motor functions, and neuronal functions of various brain parts. Anxiety tests, magnetic resonance (MR) spectroscopic evaluation, histopathological evaluation of the whole brain/specific brain parts, and brain-specific protein levels help determine brain functions.
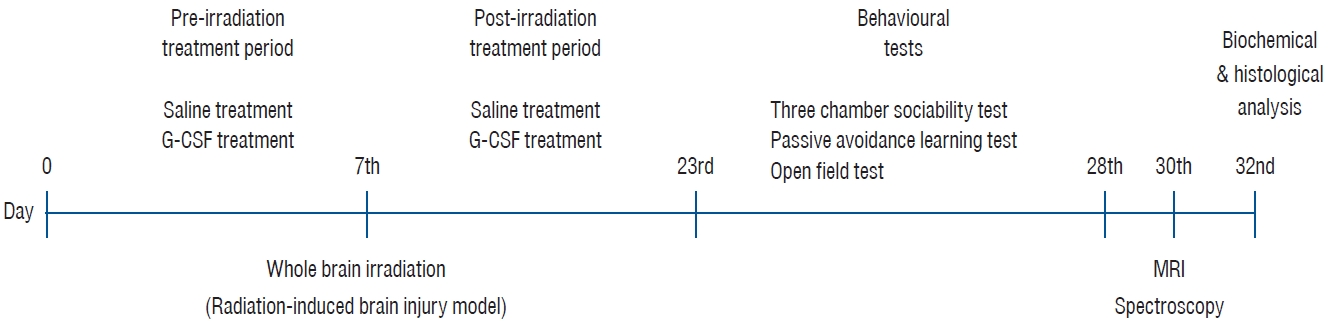
This animal model aimed to compare the rat group that received brain irradiation and did not receive additional treatment (only saline) and the rat group that underwent brain irradiation and received G-CSF treatment. In addition, the effects of G-CSF on brain functions were examined by MR imaging (MRI) and histopathologically.
MATERIALS AND METHODS
The experiments performed in this study have been carried out according to the rules in the Guide for the Care and Use of Laboratory Animals adopted by the National Institutes of Health (USA), having received the Animal Ethics Committee’s consent (Science University, Ethical number : 20220316). The rats used in the experiment were obtained from the Experimental Animal Laboratory of Science University.
Animals
This experimental study was planned by the procedures of animal studies using 24 Wistar albino female rats after obtaining permission from the Animal Ethics Committee. Ad libitum food was provided for the rats, which were kept in impenetrable steel cages with 12-hour light/dark cycles and a temperature regulated environment (22±2°C). The rats used in the study were selected at 10-12 weeks and weighed approximately 150-200 g.
Experimental procedures
In order to conduct the research, 24 female Wistar rats were acquired and split into three groups : group 1, control group who were fed orally (n=8); group 2, whole-brain radiotherapy (WBRT) with saline (n=8) : intraperitoneal administration of 0.9 NaCl saline in 1 mL/kg/day after brain irradiation; group 3, WBRT+G-CSF (n=8) : brain irradiation combined with 100 µg/kg/day of intraperitoneal G-CSF (Filgrastim; Amgen/Roche, Mannheim, Germany) (Neupogen, 48 MU/0.5 mL; Roche, Mannheim, Germany).
The eight rats (group 1) were the normal control group and had no procedure. WBRT was performed on the remaining 16 rats for the radiation injury model (groups 2 and 3). Randomly, two groups of WBRT-treated rats were created : irradiation +saline group (group 2) and irradiation+G-CSF group (group 3). Drug administration (saline or G-CSF) was started at the beginning of the study and continued for 15 days after WBRT. WBRT was given on day 7 of the start of the study. The behavioral tests were completed after 15 days. The majority of the behavioral studies were conducted during the day (from 10:00 AM to 3:00 PM). Following behavioral testing, animals were subjected to f MR spectroscopy (MRS) under anesthesia (using ketamine 50 mg/kg).
After behavioral tests and MR evaluation, all animals underwent cervical dislocation under combination anesthesia of xylazine (50 mg/kg, Rompun; Bayer, Leverkusen, Germany) and ketamine (100 mg/kg, Ketasol; Richter Pharma, Wels, Austria). All animals’ brains were removed for histopathological examination. Fig. 1 depicts the conceptual diagram for the investigation.
Irradiation procedure
Rat simulation was performed using a computed tomography scan with a 1 mm slice, and the dosage was calculated using the Eclipse treatment planning system version 8.9 (Varian Medical Systems, Palo Alto, CA, USA). Rats were anesthetized with intraperitoneal injections of 10 mg/kg xylazine and 50 mg/kg ketamine prior to irradiation. The rats were positioned on their backs. The skull was exposed to a single dose of 20 Gy of photons utilizing the source-axis distance approach with 1.0 cm of bolus material on the surface using a 6 MV linear accelerator (Clinac; Varian Medical Systems) at a dose rate of 1 Gy/min. The rats were put back in their cages after the irradiation operation. Fig. 2 depicts the WBRT apparatus and the location of the rats.
Behavioral tests
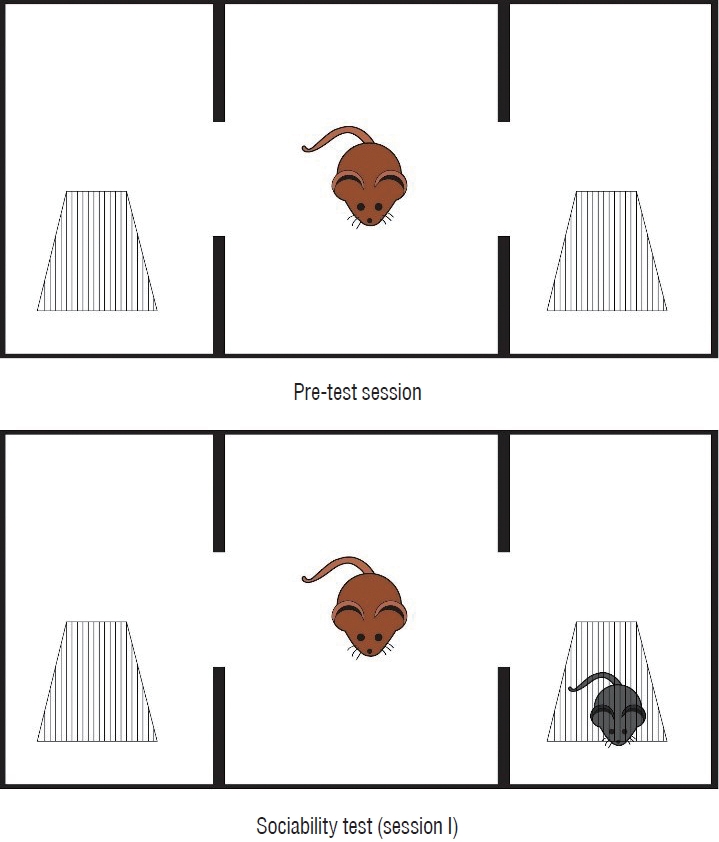
Three-chamber sociability test
This test was conducted in a 400×900×400 mm Plexiglass cage that was separated into three equal sections [10]. Rats were maintained in the test cage for 5 minutes on the first day of the pre-test period to adapt to the surroundings. Twenty-four hours later, a stranger mouse was put in a tiny plastic cage with mesh-like openings on one side and an empty cage in the third chamber to assess its friendliness. The test rat was put in the central chamber, and for 10 minutes, the duration of each zone was recorded (session 1). The test rat was deemed to have entered the chamber when its head and two front paws did so. By keeping track of the time, the proportion of time that was spent with a foreign rat was determined. To avoid olfactory stimulation, the floor was cleaned with 70% alcohol at the conclusion of each measurement. Fig. 3 depicts the threechamber sociability test prototype.
Open-field test
Two elements influence the behavioral response in this paradigm. The animal natures of rodents include the need to explore new settings (for food and refuge) and the avoidance of open and well-lit areas.
In the autism model, the open field test is believed to assist uncover stereotyped motor behavior, repetitive, automated grooming, and limitations on research activities [10]. This test is conducted in a 500 mm by 500 mm by 400 mm open-air box. At the beginning of the experiment, the rats are put in the center of the box and given 5 minutes to wander freely. After the waiting time, each rat is examined for a further 5 minutes to determine the degree of spontaneous activity. During this second 5-minute interval, the total ambulation is recorded (i.e., the number of floor segments traversed on all fours). To avoid olfactory stimulus, 70% alcohol was used to wipe the floor after each measurement.
Passive avoidance learning (PAL) test
The effectiveness of the offspring’s learning and memory is evaluated by this test [34]. It also incorporates fear-based avoidance activities where the rat learns to stay away from a safe-appearing entrance that leads to a dark room with an alarming electric grid system. The dark and light sections of the PAL box were each 200×200×200 mm in size. Rats often prefer to enter the dark chamber when put in the lit compartment. A guillotine door that divides the light and dark chamber is opened after a 10-second acclimation period. The door between the light and dark areas is shut when the test rat enters the dark area. The experimental rat receives a 1.5 mA electric shock in the dark for 3 seconds, after which it is removed and put back in its cage. The mice are put back inside the PAL box after a 24-hour waiting period. This time, no shock is administered; instead, the duration of the rat’s passage from the light to the darkroom is measured or recorded. The delay time is measured up to a maximum of 300 seconds, and the amount of time it takes the rat to decide not to enter the dark room is inadvertently used as a memory indicator.
MRI protocol and MRS
Conventional MRI
Each rat was evaluated using a 3.0-T MRI/MRS scanner (Magnetom; Siemens Healthcare, Erlangen, Germany). Typical MR sequences consisted of the following : sagittal fast spin-echo T1 weighted imaging for location; axial spin-echo T1 weighted imaging (repetition time [TR] / echo time [TE] = 400/11 ms; field of view [FOV] = 60 mm; matrix = 256×256; the number of excitation pulses = 2; bandwidth = 12.5 kHz; slice thickness = 1 mm; interslice gap = 0.2 mm; the total number of scan slices = 16); fast spin-echo T2 weighted imaging (TR/TE = 4000/120 ms; other parameters identical to those used in the T1 weighted spin-echo sequence); and fast fluid attenuation inversion recovery (TR/TE = 4000/120 ms; time of inversion = 2200 ms; matrix = 256×192; the number of excitation pulses = 1; other parameters identical to those used in the T1 weighted spin-echo sequence).
MRS
In order to perform automated multivoxel 2D chemical shift imaging on all rats, the following parameters were used : TR = 1000 ms; TE = 35 ms; phase encoding x = 24; phase encoding y = 24; and the number of excitation pulses = 1. Slice thickness was 4 mm, the FOV was 60 mm in diameter, and the MRS’s voxel size was 1.87×1.87×4 mm3. Right striatum was the area of interest that was selected. Five hundred eighty seconds were spent on the collection of the 2D 1H-MR spectrum in total. The raw data kept at a workstation was processed by Magnetom software (Siemens Healthcare).
Histopathological evaluation of hippocampus and cerebellum
The cornu Ammonis (CA) 1 and CA3 areas of the hippocampus were used to assess hippocampal injury. On brain slices, cerebellar damage was also evaluated. The animals underwent predetermined behavioral tests before being put to death. Brains of animals were removed. Fixation was place for three days using 10% formaldehyde in 0.1 M phosphate buffer saline (PBS). After that, the materials were kept in a 30% sucrose solution at 4°C until the infiltration was finished. Following that, coronal sections of each brain were removed and put on gelatinized glass slides using a microtome set to a 40 µm sliding distance.
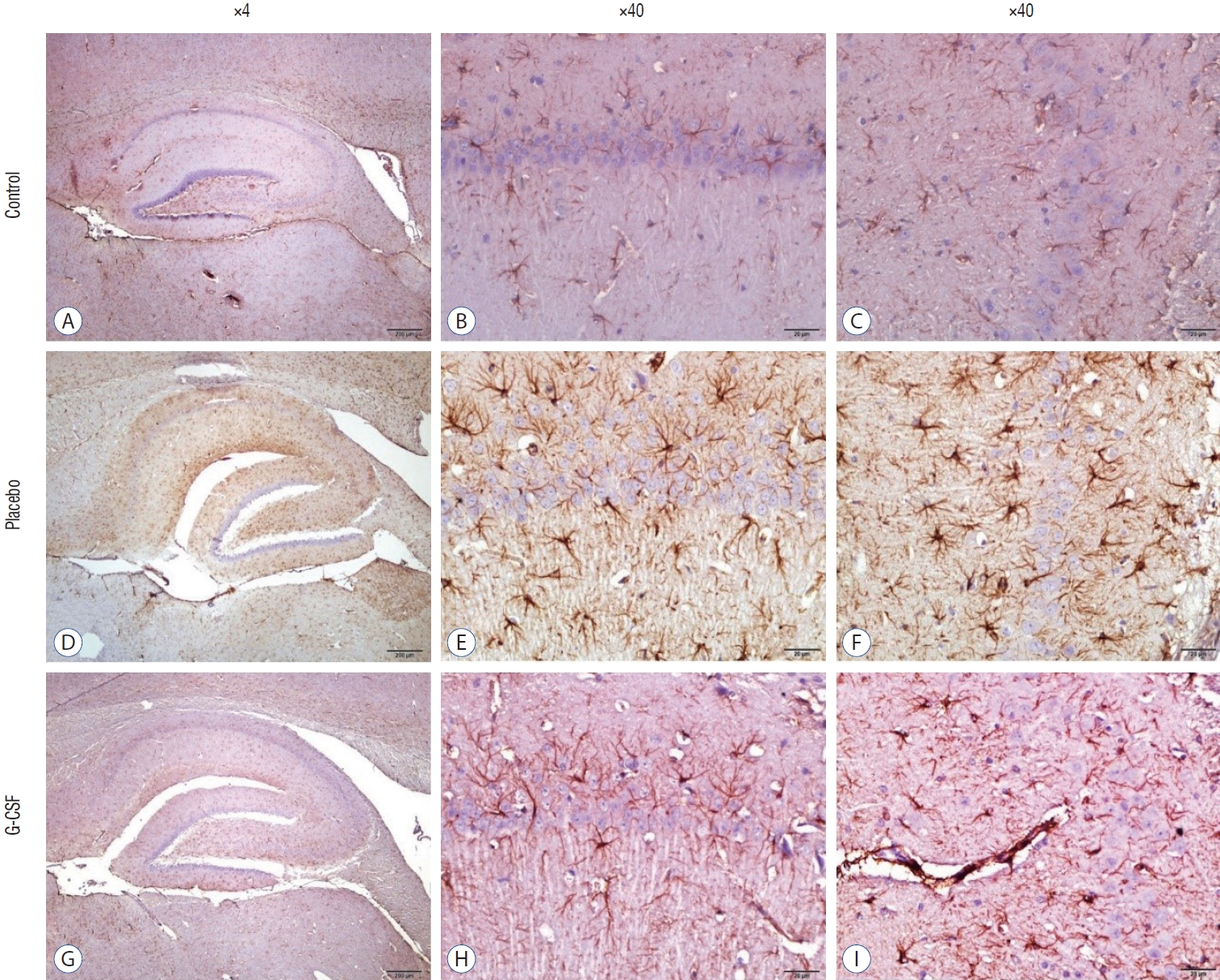
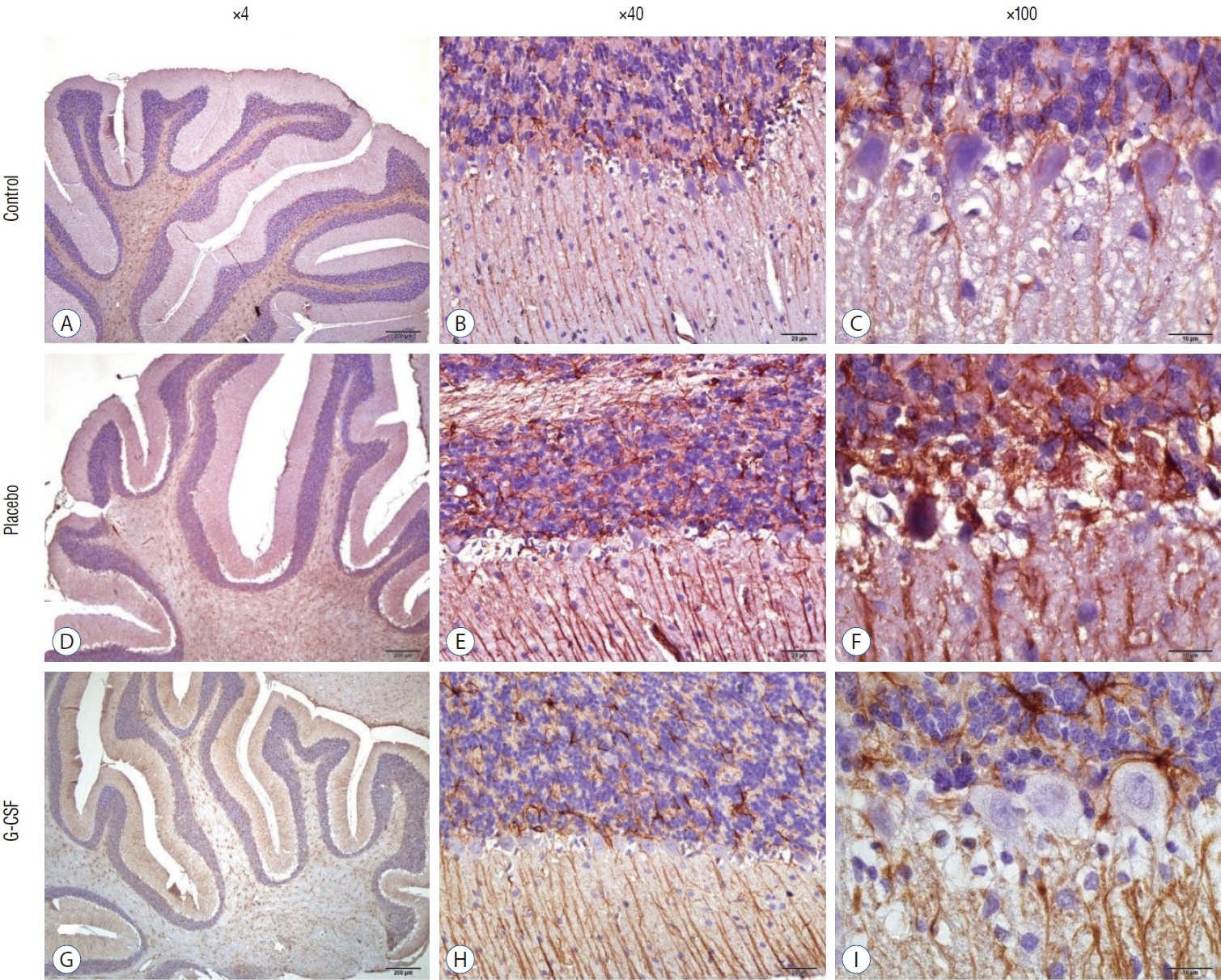
With the use of a Cresyl Violet Acetate solution, Nissl material was found in the cytoplasm of neurons, and the number of surviving neurons was counted for each research group. On the other hand, for glial fibrillary acidic protein (GFAP) immunohistochemistry, brain slices were pre-treated with 10% H2O2 for 30 minutes and then blocked with 10% normal goat serum (Invitrogen, Carlsbad, CA, USA) for an hour at room temperature. Sections were incubated with primary GFAP antibodies for 24 hours at 4°C (Abcam, Inc., Cambridge, MA, USA; 1/1000). Antibody detection was carried out using the Histostain-Plus Bulk kit from Invitrogen against rabbit immunoglobulin G. Following 3,3’-diaminobenzidine (DAB) viewing of the finished product, the sections were cleaned with PBS. In random sections, the number of GFAP-positive cells was counted at ×40 magnification. The same scientist who was blind to the research groups conducted each histopathological investigation.
Brain biochemical analysis
The brain tissue that was taken during the decapitation procedure was kept at a temperature of -20°C until the research period. After being thoroughly homogenized in 5 L of phosphate-buffered saline with a pH of 7.4, all brain tissues were subsequently centrifuged at 5000 g for 15 minutes to remove any remaining debris. Following the collection of the supernatant, the total protein content in the brain homogenates was assessed using Bradford’s technique and a reference standard of bovine serum albumin.
Using commercially available rat enzyme-linked immunosorbent assay kits, the levels of malondialdehyde (MDA), brain-derived neurotrophic factor (BDNF), NF-κB, tumor necrosis factor-alpha (TNF-α), postsynaptic density protein-95 (PSD-95), and lactate were assessed in the brain supernatants. The absorbances were measured using a microplate reader (MultiscanGo; Thermo Fisher Scientific Laboratory Equipment, Waltham, MA, USA).
Statistical analysis
For statistical analysis, SPSS version 23.0 for Windows (SPSS Inc., Chicago, IL, USA) was utilized. Levene’s test was used to determine homogeneity of variance while the Shapiro-Wilk test was used to determine normality. Mean and standard error of the mean are used to represent the findings (standard error of the mean). Multiple groups were compared using a one-way analysis of variance test. It was agreed that the result of p<0.05 was statistically significant.
RESULTS
The comparison of behavior tests in groups is shown in Table 1. The average amount of time spent with the new rat was 65.6±4.8 in group 1, 29.5±4.4 in group 2, and 60.7±3.2 in group 3. G-CSF treatment significantly increased the sociability test results. In addition to the sociability test, the open field test and PAL test showed better results with the G-CSF treatment.
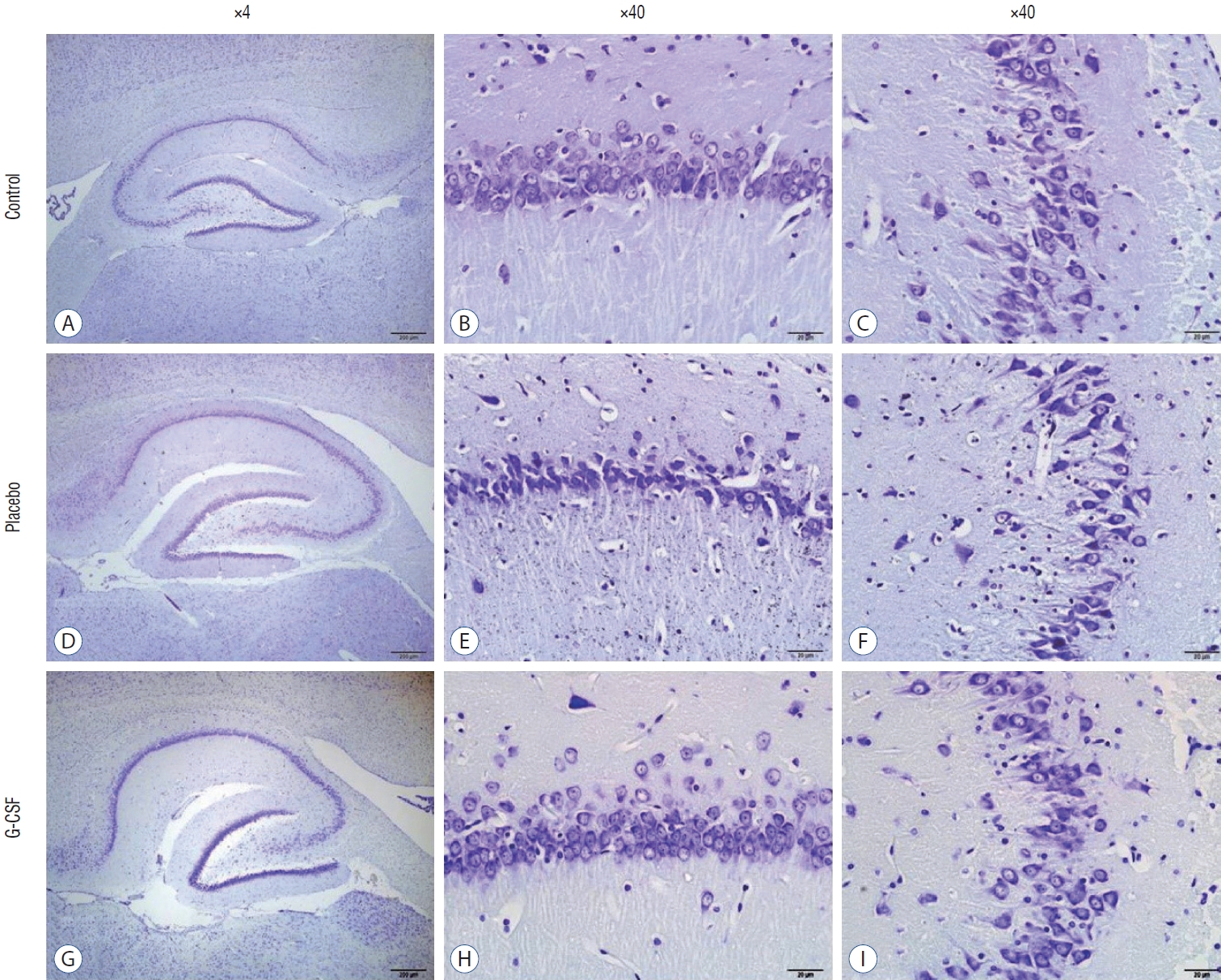
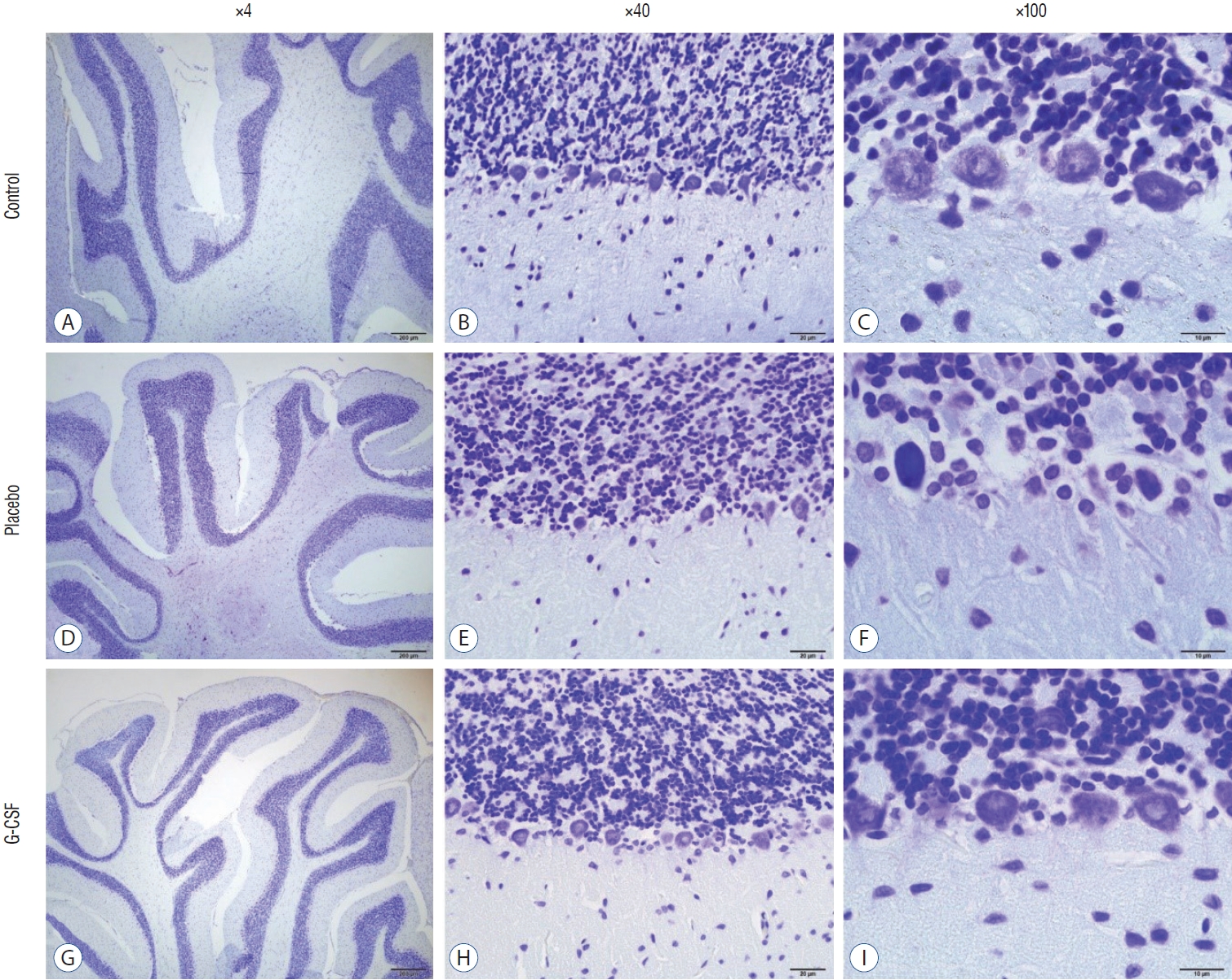
The counting of CA1, CA3, and Purkinje neurons were used in hippocampal and cerebellar neuron evaluation. In addition, all neurons’ GFAP immunostaining indexes were calculated. The comparison of neuronal counts and GFAP immunostaining indexes in groups are shown in Table 2. All neuron counts decreased after brain irradiation, but all GFAP immunostaining indexes increased. However, while the mean number of neurons in group 3 was statistically higher than in group 2, the mean of group 3 GFAP immunostaining index was statistically lower than group 2. CA1 and CA3 neurons (Fig. 4) and Purkinje neurons (Fig. 5) in the brains of female rats in the G-CSF group show enhanced neural morphological modifications and increased neural count (Fig. 4). However, brain irradiation and G-CSF female rats have decreased glial activity in CA1 and CA3 areas (Fig. 6) and Purkinje neurons (Fig. 7).
The mean levels of MDA, TNF-α, NF-κB, and lactate in group 2 were higher than in group 1, while BDNF and PSD-95 levels of group 1 were higher in group 2. After G-CSF treatment, a decrease happened at the mean levels of MDA, TNF-α, NF-κB, and lactate, and an increase occurred at the mean levels of BDNF and PSD-95. The comparison of brain protein levels in groups is shown in Table 3.
The comparison of the brain lactate levels in groups is shown in Table 4. The mean brain lactate level increased after brain irradiation, but the mean lactate level decreased with G-CSF treatment. On MRS, the high lactate peak is seen, and regular peaks are not seen due to brain irradiation injury. After G-CSF treatment, typical peaks of brain metabolites started to be seen, and the high lactate peak had lost its level. MR spectroscopic evaluation of brain metabolites is shown in Fig. 8.
DISCUSSION
Radiotherapy is a treatment modality for primary CNS tumors and metastases [29]. While radiation therapy affects tumor cells, it also affects normal brain cells. The effects of cranial radiotherapy on cognitive functions determine the quality of life. Especially in treating brain tumors, quality of life is as important or should be as important as living a long time. Radiation can cause impairment of functions such as thought, memory, concentration, and speech and reduce the quality of life [17]. Diffuse leukoencephalopathy and cerebral atrophy can also develop with the effect of cranial radiotherapy [37]. According to the results of this animal brain irradiation model, it was first observed that G-CSF had reparative, neuroprotective and anti-neurodegenerative effects and had increased neurotrophic factor expression in the brain irradiation animal model.
Using hematopoietic growth factors such as G-CFS in treating some brain injuries has been groundbreaking [31,33]. In the research by Song et al. [35], treatment of G-CSF for 3 days after a moderate traumatic brain injury led to recovery of performance in a water maze, an assessment of memory and learning, 2 weeks after the trauma. Moreover, previous studies showed that intraperitoneal or intravenous administration of G-CSF had positive effects in animal models [1,40]. Various studies have shown that single G-CSF treatment modalities can also be used in traumatic brain injury for cognitive function recovery [32] or cellular proliferation with motor function recovery [27]. The combined treatment of G-CSF and human umbilical cord blood cells, on the other hand, was shown by Dela Peña et al. [7] to lessen neuroinflammation in traumatic injury of brain. In an animal model of chronic traumatic brain injury, Acosta et al. [1] demonstrated that a combined treatment of human umbilical cord blood cells and G-CSF improved histopathological and motor deficits. This study showed that single-use G-CSF treatment in brain irradiation positively affected animals’ brains.
ROS are delicately regulated in brain tissue under physiological settings by the interaction of pro-oxidant and antioxidant mechanisms. Still, in the case of brain injury, an increase in the production of ROS occurs [38]. MDA, an end product created during lipid peroxidation, is likewise increased by brain injury since it also induces lipid peroxidation. In many studies conducted in the past, it has been shown that the MDA level is increased, especially in trauma cases with a mortal course [21]. On the other hand, animal studies found that the antioxidant agents reduced MDA levels in brain tissues [12,14]. In the present study, while an increase was observed in the mean MDA levels in the rat groups in which brain irradiation was applied (from 42.9±2.4 nmol/g to 83.1±6.5 nmol/g), it was observed that the mean MDA level was decreased up to 69.3±7.7 nmol/g in the G-CSF applied group compared to the other groups.
Cytokines are proteins produced by neurons and glial cells during brain inflammation that modulate the inflammatory process. Expression of TNF-α secreted by neurons and glial cells accelerates the movement of inflammatory cells to the site of inflammation. In a previous study, experimental stroke was created with permanent middle cerebral artery occlusion in rats; it was found that TNF-α mRNA expression peaked at 12 hours [16]. Another clinical study showed that TNF-α levels increased in the cerebrospinal fluid in febrile epilepsy [28]. This animal model’s mean TNF-α level increased significantly due to irradiation. Still, the mean TNF-α level was considerably lower in the group (52.3±5.4 pg/mg) receiving G-CSF compared to the brain irradiation+saline group (128.1±9.01 pg/mg).
In the latter part of the 20th century, the NF-κB transcription factor was identified [2]. Later research showed that it is present in almost all of the organism’s cells, despite its existence having previously been shown in immune cells. NF-κB, which activates roughly 500 genes, is important in processes including immune response, cell cycle, survival, proliferation and development, plasticity, and memory formation [30]. By effectively supplying long-term potentiation, NF-κB influences the CNS’s plasticity and, therefore, memory formation. Recent years have seen a rise in research on the significance of inflammation’s role in neurodegeneration. One of the key processes of inflammation involves NF-κB in a crucial way [39]. In this animal model, the mean level of NF-κB protein increased to 115.6±8.5 pg/mg due to brain irradiation, which was significantly higher than the control group. However, adding G-CSF treatment to brain irradiation lowered the mean NF-κB protein level to 45.7±3.9 pg/mg.
Lactate is released from the brain due to brain hypoperfusion [6]. After brain injury, due to mitochondrial dysfunction, pyruvate to the tricarboxylic acid cycle increases and is converted to pyruvate lactate by the enzyme lactate dehydrogenase. Therefore, lactate level increases significantly in cases of brain injury. Dumbuya et al. [8] showed that G-CSF had a crucial neuroprotective role in neonatal hypoxic-ischemic encephalopathy. In the present animal model, the mean brain lactate level decreased with G-CSF treatment significantly.
One of the neurotrophic factors, BDNF, is crucial for the differentiation, maturity, and survival of neurons that operate as neuroprotective agents in dangerous situations by promoting and regulating neurogenesis [4]. In the work by Song et al. [35], it was shown that mice treated with G-CSF boosted their levels of BDNF produced by activated microglia and astrocytes in an animal model of traumatic damage. Additionally, G-CSF treatment 3 days after moderate traumatic brain damage led to the recovery of performance in a water maze, a test of memory and learning, 2 weeks following injury [35]. The mean BDNF level rose with G-CSF therapy in this animal model compared to the brain irradiation+saline group, which is consistent with the literature.
A postsynaptic scaffolding protein in excitatory neurons is called PSD-95. It has previously been shown that G-CSF greatly boosted the cell survival rate of hippocampal neurons in vitro and raised the expression of PSD-95 in the hippocampus [15]. It was established in the present model that the mean level of brain PSD-95 protein was 0.93±0.04 pg/mg, which decreased to 0.36±0.15 pg/mg after brain irradiation. However, the PSD-95 protein level increased significantly to 0.67±0.22 pg/mg due to the G-CSF application.
Many tests, such as sociability tests, open field tests, elevated plus/zero maze interaction tests, light-dark tests, hole board tests, and ladder tests, can be used to evaluate anxiety in studies on experimental animals. There are three phases to the exam. Stage 1 entails becoming acquainted with the apparatus; stage 2 requires putting a stimulating animal into one side chamber; stage 3 is optional and involves placing a second stimulating animal in the opposing side chamber [18]. Mice and rats tend to spend more time with the new animal with an innate curiosity. Previous animal models showed reduced social recognition memory and sociability after experimental brain injuries in rats [19]. Loss of hippocampal volume and decreased neuroplasticity in the hippocampus and prefrontal cortex have been linked to abnormalities in social functioning following experimental brain traumas, however this association has not yet been confirmed [3,20]. In the presented study, only stages 1 and 2 were tested. According to a three-chamber test, the mean value of sociability percent decreased after brain irradiation (from 65.6% to 29.5%) but was re-gained after G-CSF treatment (from 29.5% to 60.7%).
Open-field tests are used to measure exploratory locomotor activity since they are based on an animal’s intrinsic desire to explore a new habitat. Open spaces emphasize the intrinsic aspects of exploratory behavior, while motor impairments and cognitive dysfunction greatly influence this behavior. In previous studies, animals may exhibit decreased activity, regular activity, or hyperactivity in the open field depending on the severity and form of the brain injury [26,36]. In this animal model, irradiated rats showed hypoactive ambulation, whereas irradiated G-CSF-administered rats showed decreased ambulation compared to the control group but increased compared to the irradiation+saline group.
PAL is a hippocampus-dependent behavior, and it has been shown that the delay time to enter the darkroom is reduced in animal models with hippocampal lesions [11]. Our animal model showed that PAL was lower due to brain irradiation, and improved PAL was observed with G-CSF treatment applied during brain irradiation.
The hippocampus is a grey matter layer located in the medial temporal lobe. It plays a vital role in memory, emotion, positioning, and direction finding, and it forms the limbic system together with the cingulate gyrus, hypothalamus, and amygdala [13]. Two regions at the hippocampus (CA1 and CA3) indicate the development of spatial learning and memory. On the other hand, Purkinje neurons are the primary cells of the cerebellum. Previous studies showed that brain injuries resulted in cognitive impairments. They reduced the number of CA1, and CA3 neurons [5,9]. In the present study, CA1, CA2, and Purkinje neuron counts decreased due to brain irradiation, but these neuron counts increased after G-CSF treatment.
A marker for brain damage is called GFAP, a significant intermediate filament [41]. After a brain injury, astrogliosis (or reactive astrocytosis) is a critical process for healing the injured brain area. In a spinal cord injury model of rats, concomitant G-CSF treatment and exercise therapy are effective for motor function recovery [22]. In the current animal model, GFAP immunostaining indexes of CA1, CA3, and Purkinje neurons in the brain irradiation-G-CSF group were lower than in the brain irradiation-saline group.
Excessive production of lactate caused by glycolysis in the brain is considered an indicator of altered metabolism. It indirectly indicates an ischemic change in the brain parenchyma with insufficient perfusion or increased necrotic tissue due to irradiation and edema. Mitochondrial diseases with cerebral involvement, hypoxia/ischemia, inflammation, and tumors are the etiological diseases that cause increased lactate levels [25]. Due to the hypoxic/ischemic and inflammatory effects of brain irradiation, the mean brain lactate level was higher than the control group (100 vs. 518.6), but adding G-CSF to brain irradiation had a lactate-lowering effect.
CONCLUSION
Radiotherapy is a treatment modality for primary central nervous system tumors and metastases. While radiation therapy affects tumor cells, it also affects normal brain cells. On the other hand, diffuse leukoencephalopathy and cerebral atrophy can also develop with the effect of cranial radiotherapy. The positive impact of growth factors has been demonstrated in animal models of traumatic brain injury previously, and this study was designed with the conclusion that they may have similar effects. According to the results of this animal model, it was first observed that G-CSF had reparative, neuroprotective and anti-neurodegenerative effects and had increased neurotrophic factor expression, neuronal counts, and morphology changes. In addition, G-CSF had a proven lactate-lowering effect in MRS and brain materials.