INTRODUCTION
Intracranial aneurysm (IA) refers to a bulge in the wall of intracranial arteries due to endothelial dysfunction and extracellular matrix remodeling of the hemodynamic response. Although the prevalence of IA is approximately 3% in the general population, the mortality rate due to subarachnoid hemorrhage following aneurysm rupture is close to 50% [25,31]. Clinical and radiological studies showed that IA formation and rupture were related to female gender, hypertension, smoking, larger size and posterior circulation aneurysm [3].
The plethora of genome-wide association studies (GWASs) during late 2000’s has increased the number of investigations into IA. These genetic studies have identified several candidate genes and loci associated with IA such as EDNRA, GBA, CDKN2A/B, RBBP8, STARD13/KL, and SOX17 [1,2,11,29]. GWAS reported differences in the frequency of single-nucleotide polymorphisms (SNPs) based on case-control studies. Accordingly, the missing heritability is explained by independent SNPs involving complex diseases in human including cerebrovascular diseases (CVDs). To address this issue, robust analytical approaches such as meta-analyses, multifactorial interactions, and polygenic risk scoring systems have been performed in many populations [9]. A GWA meta-analysis reported IA-associated risk loci, including new loci (SLC22A5, 6q16.1, 12q21.33, PSMA4, and NT5C2) based on 10754 cases and 306882 controls [1]. Two large-scale meta-analyses also reported the successful validation of two IA-associated loci boule (BOLL) and endothelin receptor type A (EDNRA) in an East Asian population of nearly 20000 individuals [12,13]. However, few studies have reported gene-gene interactions or polygenic risk scores for IA patients. Considering to two previous findings of BOLL and EDNRA loci, here we estimated the effects of genome-wide gene-gene interactions on IA in a prospective hospital-based cohort study.
MATERIALS AND METHODS
All the study protocols have been approved by the Institutional Review Board and Ethics Committee of Hallym University Chuncheon Sacred Heart Hospital (No. 2016-3, 2019-06-006). The study protocol and design are described in detail elsewhere [11].
Study populations
The study subjects were enrolled from the multi-institutional biobanks comprising five university hospitals constituting “The First Korean Stroke Genetics Association Research”, and including patients diagnosed with CVDs such as IA between March 2015 and December 2020 (https://www.1ksgh.org/) [11,18]. Data derived from 250 patients with IA and 296 controls, which were also used in the first Korean IA GWAS [11], were used in the analysis. The inclusion of patients with IA was based on the following criteria : 1) adult patients more than 18 years of age; 2) patients without other types of CVD such as ischemic stroke, hemorrhagic stroke, and vascular malformation; and 3) patients without any other genetic disorders such as polycystic kidney and moyamoya disease. Control subjects were defined as adults without CVD. Medical and radiological data were collected and updated.
Genotyping and quality controls
Genomic DNA derived from the peripheral blood of the study population was genotyped using the AxiomTH Asian Precision Medicine Research Array (APMRA) (Thermo Fisher Scientific, Waltham, MA, USA). High quality plates were defined by a plate pass rate higher than 95% for samples and the average call rate of passing samples was greater than 99%. Out of 798148 SNPs, 512575 SNPs passed the quality control including genotyping call rate of 95% or higher, minor allele frequency (MAF) of at least 1%, and Hardy-Weinberg equilibrium (HWE) with p-value ≥1×10-6 [11].
Genome-wide SNP-SNP interactions via BOLL and EDNRA genes
We investigated genome-wide SNP-SNP interactions using either BOLL or EDNRA loci associated with IA in previous GWASs and meta-analyses [11-13]. The multivariate analyses of the SNP interactions between rs700651 intron SNP (BOLL, 2q33.1) and rs6841581 upstream SNP (EDNRA, 4q31.22) on 512574 SNPs were performed using the Contrived Acronym of software for SNP Interactions (CASSI ver. 2.5; https://www.staff.ncl.ac.uk/richard.howey/cassi/index.html) [27]. SNP-SNP interactions were analyzed by choosing target SNP from two given SNP windows (possibly from different pedigree files). Each pair of SNPs that interaction test passes a given significance level (i.e., minimized p-value=1) is returned in the output file with possible extra information such as beta coefficient, standard error, chi-square, and p-value. The CASSI accepted only PLINK binary files in order to perform the calculations as efficiently as possible. The logistic regression epistasis test was available for SNP-SNP interactions in this study even though this program can provide the maximum number of 1M terms. Subsequent regression analyses were carried out under the additive effect model and adjusted for 10 covariates including age, sex, hypertension, diabetes mellitus, hyperlipidemia, smoking status, and four genetic ancestry factors. The effect coefficient was estimated using the natural log-transformed scale of odds ratio (i.e., natural log-transformed odds ratio [lnOR]). The multiple comparisons were adjusted for a genome-wide interaction threshold with a p-value less than 1.25×10-8 (genome-wide p-value=5×10-8 divided by four tests for interaction). A subsequent power and sample size calculations for each interaction term were estimated under the assumption with GW interaction significance threshold, 1 : 1.2 case-control ratio and information of each SNP (i.e., allele frequency and effect size) through performing the Quanto ver. 1.2.4 program (https://bio.tools/QUANTO). The performance of the large-scale interaction analyses was completed by the multi-tasking supper computer that has a capacities of Intel(R) Xeon(R) CPU E5-2667 v4 (3.20 GHz), 256 GB RAM, and 15 Cores. We performed Manhattan plots of BOLL and EDNRA interaction using the package of “qqman” in R v3.6.1 (https://cran.r-project.org/web/packages/qqman) and regional visualizations of the target SNP’s base-pair position ±400 kb regions using LocusZoom ver. 1.3 (https://genome.sph.umich.edu/wiki/LocusZoom_Standalone) written in Python and R [23].
RESULTS
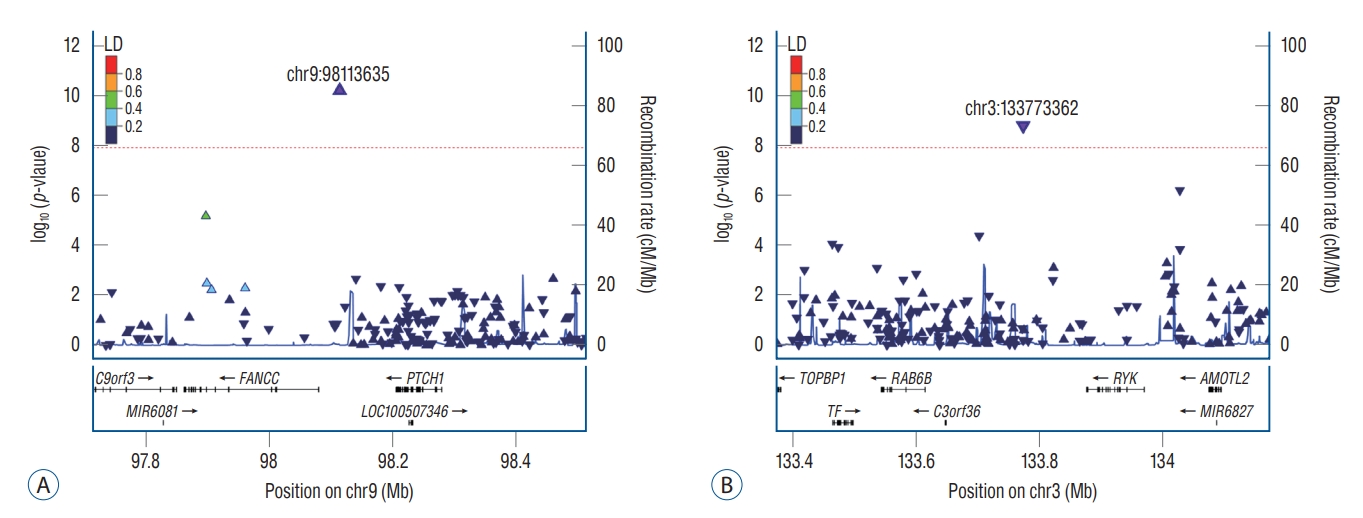
Detailed information including SNP genotype distribution and HWE p-value, is presented in Supplementary Table 1. Out of 512574 SNP interaction terms, 23 and 11 SNPs reached a genome-wide interaction threshold (p<1.25×10-8) with rs700651 intron SNP (BOLL, 2q33.1) and rs6841581 upstream SNP (EDNRA, 4q31.22), respectively (Fig. 1, Tables 1 and 2). All 34 SNPs showed an MAF above 1% and an HWE p-value greater than 0.01 (Supplementary Table 2). Most of the BOLLor EDNRA-interacting SNPs showed shared alleles in both patient and control groups (i.e., average of MAF >0.23) without significant association in a single SNP analysis (0.0<p<1) (Table 1). These findings suggest that BOLL or EDNRA may have a synergistic effect on IA formation, via multifactorial pair-wise interactions, rather than involved alone in the IA formation. Among the interactions, the rs1105980 upstream SNP of PTCH1 gene (9q22.32) showed the most significant interaction with rs700651 (effect, 1.53; p=6.41×10-11) (Fig. 2A). The rs74585958 of RYK gene (3q22.2) interacted strongly with rs6841581 (effect, -19.91; p=1.64×10-9) (Fig. 2B). Two strong pair-wise linkage disequilibrium (r2>0.95) were observed in the interaction of rs328025 with rs700855 (RGPD4, 2q12.3) and between rs11925024 and rs1231 (TNIK, 3q26.31) (r2>0.95, data not shown). Interestingly, two EDNRA-interacting gene variants of TNIK (rs11925024 and rs1231, p=1.04×10-8 and 1.22×10-9, respectively) and FTO (rs9302654, p=3.78×10-9), and one BOLL-interacting METTL4 gene variant (rs549315, p=4.80×10-10) showed marginal interaction with BOLL gene (0.001<p<0.05). However, there was no direct interaction between BOLL and EDNRA variants (effect, -0.27; p=0.301). When power calculation was estimated by the basis of detail SNP information (Supplementary Tables 2 and 3), two SNPs such as rs74585958 (RYK) and rs150664966 (EIF4H) showed sufficient statistical power among EDNRA-interacting loci (i.e., 0.885 and 0.805, respectively). The rs11672303 (SLC7A10) of EDNRA-interacting loci showed a marginal statistical power of 78.9%. In contrast, no SNP reached sufficient statistical power threshold of 80% in interaction terms by the BOLL locus (i.e., power <50%).
DISCUSSION
Although BOLL gene, the G allele of rs700651 associated with risk showed a significant genome-wide overall effect on IA in multi-ethnic integrative meta-analyses (p=1.05×10-8) [12], its pathgeneic mechanism remains to be identified. Most studies related to BOLL involved spermatogenesis due to its role in germ cell development or cancer [17,22]. BOLL is a well-known gene associated with normal germ cell development [26]. The gene is predominantly expressed in secondary spermatocytes [28]. However, few studies investigated the role of BOLL in CVD including IA. Harrod et al. [10] reported that estrogen deficiencies may lead to IA by interrupting the inflammatory response. In reality, earlier age at menopause increased the risk of IA, suggesting the association between estrogen deficiency and IA pathogenesis [6]. Thus, in the case of BOLL gene, additional studies are needed to determine the protective effect of male hormones against the IA development. The EIF2B5 gene that interacts with BOLL in the current GW interaction and network analyses exhibited a homologous inhibition of cell translation. The differential expression of EIF2B5 was moderate in human tissues and cells. Brady et al. [4] reported that intron retention in EIF2B5 inhibited protein translation in hypoxic cancer cells. In the case of abdominal aortic aneurysms, there was no meaningful EIF2B5 network [16]. Based on a review of the current literature, it is difficult to elucidate the contribution of the two genes to IA pathogenesis. Thus, a further in vivo study is needed to investigate the function of the two genes in IA formation.
The G allele of the rs6841581 located near the 5’-untranslated region of EDNRA (4q31.22) gene was associated with IA [13]. However, detailed mechanisms of IA mediated by EDNRA have yet to be reported to determine the direct effect or an indirect effect. Rats with pulmonary hypertension showed higher expression of EDNRA genes [21]. Endothelial injury, followed by disruption of collagen and elastin synthesis contribute to IA [15]. Chronic hypertension per se may induce the structural changes. Thus, the inflammtory response to increased hemodynamic stress following the disruption of cerebral arteries mediated by EDNRA may result in IA. In our study, among the EDNRA-interacting genes, the MPDZ-centered interaction between EIF4H, SAP18, and UNC13C was observed. MPDZ is a tight junction protein, which modulates notch signaling during angiogenesis by controlling ligand recruitment to adherent junctions [8,24]. Feldner et al. [8] reported that loss of MPDZ decreased ependymal cell integrity and caused hydrocephalus. Ependymal cells are mainly responsible for electrolyte transport between brain parenchyma and the CSF. Adult ependymal cells are highly differentiated. Ependymal cells lining the lateral ventricles are quiescent under normal physiological conditions [5]. However, after stroke, adult ependymal cells are transformed into radial glial cells in the subventricular zone [30]. IA formation and growth occur within the CSF space surrounding the cerebral arteries. Accordingly, a further study is required to investigate the relationship between IA and CSF mediated by ependymal cells.
In our study, most variants did not show a significant association with IA via single SNP-based GWAS, although they exhibited significant associations with BOLL or EDNRA via multiple interactions terms. It is widely believed that a single SNP often has small effect on disease phenotypes including IA, thus it cannot fully account for the genetic susceptibility, in particular stroke. Therefore, identification of SNP interactions that are associated with disease is increasing to interpret the genetic basis of the disease susceptibility [20]. Although several loci related to BOLL or EDNRA did not pass genome-wide significance in a single SNP analysis, the loci might have a synergy effect on IA development by interacting with the genes. In this study, we aimed to investigate gene-gene interaction using the previous GWAS data for Korean patients with IA for the first time [11]. However, due to the relative small number of the enrolled patients and possible false positives [14], we did not perform all possible pairwise SNP-SNP interaction and inevitably focused on two IA candidates of BOLL and EDNRA by referring to previous studies [12,13]. Nevertheless, we required further replication GWAS and exhaustive searching for SNP-SNP interaction in a large dataset of GWAS [19]. In addition, further molecular functional study including the estimation of protein levels to validate our interaction results between BOLL (or EDNRA) and several loci in the future.
The study has some limitations. First, although we identified novel loci for IA, their functional role was not investigated. The role of most of the 34 novel genes interacting with BOLL and EDNRA in our study has yet to be analyzed in IA or other CVDs. Second, the study had a potential sample size limitation of multiple interaction terms with either BOLL or EDNRA loci, which have been replicated in IA susceptibility involving Korean adults including 250 patients with IA and 296 controls. In addition, there is a possibility that the results of this IA genetic study are limited to the Korean population. Therefore, we may warrant these findings in the second stage GWA meta-analysis and interaction test. Nevertheless, this study evaluates the first multiple genome-wide SNP-SNP interactions by IA-targeting genes such as BOLL and ENDRA, which have been validated in previous studies [12,13]. Disease is caused by various biological pathways and it is difficult to explain it based on a GWAS, which focused on differences between individual paired loci [7]. Accordingly, a study of genetic interactions and additional insights into various compensatory functional modules is needed to elucidate complex diseases such as IA. In summary, it is necessary to develop a general framework for mapping complex genetic networks of IA using GWAS data combined with clinically relevant risk factors.
CONCLUSION
Genome-wide interaction between IA and BOLL or EDNRA revealed 34 novel loci, which were likely to be associated with IA. Common susceptibility variants and their interacting factors can be used to determine the inter-individual status of IA formation. The novel gene-gene interactions reported in this study need to be corroborated via larger prospective cohort studies.