INTRODUCTION
Spinal cord injury (SCI) results in neurological deficits in both sensory and motor functions, leading to inevitable disabilities [14]. Currently, there is no available treatment that can revert to an intact state. In a previous study, we focused on axonal regeneration for repair of SCI, and evaluated the efficacy of Wnt3a-secreting cell transplants in regenerating injured axons; we observed increased axonal regeneration and recovery of motor function in a sub-acute SCI [21,25,26]. Wnt proteins are a family of signaling molecules that play roles in development, cell differentiation, and proliferation [8,23]. They are also crucial in multifunctional axon guidance [8,28]. Wnt signaling through glycogen synthase kinase-3 is involved in axon remodeling [8,28]. Upregulation of Wnt signaling activates Ryk, a Wnt receptor in corticospinal neurons, resulting in axon retraction after injury [18]. Wnt1 and Wnt3a modulate spinal cord dorsal interneuron differentiation, and Wnt3a has been implicated in neurogenesis [7].
Currently, high proportion of candidates for cell therapy in SCI is chronic status, including those in our previous clinical trials [16,20]. However, there is no gold standard for determining the chronic phase period in a rat model of SCI. To standardize a practical approach to chronic SCI, we need to identify the most suitable chronic phase in a rat model of SCI. In our pilot study, we injected mesenchymal stem cells (MSC) at 4 weeks after SCI, but found that chronic SCI manifests 6 weeks after SCI; behavioral improvement plateaued at 6 weeks after SCI in most injury only group (data not shown). Therefore, in this study, we transplanted Wnt3a-secreting human mesenchymal stem cells (hMSC) into a contusion site 6 weeks after SCI. We also hypothesized that enhanced Wnt3a protein expression could induce more pronounced neuro-regenerative effects even in the chronic condition. Thus, this study aimed to identify the ideal expression ratio of the Wnt3a gene transfected into MSC to maximize neuro-regenerative effects in a chronic rat SCI model.
MATERIALS AND METHODS
All procedures complied with the guidelines of the Institutional Animal Care and Use Committee of the Asan Institute for Life Sciences and were approved by the Ethics Committee for Animal Experiments of the Asan Institute for Life Sciences (Seoul, Korea).
Cloning and lentivirus production
We extracted human Wnt3a cDNA from a pcDNA/Wnt3a plasmid (#35908; Addgene, Cambridge, MA, USA) using restriction enzymes. We inserted extracted DNA into a pLentisuCMV-Rsv-GFP plasmid (GenTarget Inc., San Diego, CA, USA) to create a pLenti-suCMV-Rsv-GFP/human Wnt3a plasmid. For lentivirus production, we used a Neon transfection System (Thermo Fisher Scientific, Waltham, MA, USA), mixing pLenti-suCMV-Rsv-GFP/human Wnt3a, pMD2G (#12259; Addgene), and psAX2 (#12260; Addgene) in Lenti-X 293T (Takara Bio, Shiga, Japan) cells. After 2 days of incubation, we harvested the supernatant containing the concentrated virus using the WelProt Virus Concentration Reagent (WelGENE, Daegu, Korea). Lentivirus titers were measured using the Lenti-X qRT-PCR Titration kit (Takara Bio).
Genetically modified MSC
MSCs derived from human umbilical cord blood (KS-R218-3G1-N; Kangstem Biotech, Seoul, Korea) were multiplied and cultivated in a KSB-3 Complete Medium® kit (Kangstem Biotech) at 37ºC and 5% CO2. These cells were infected with the prepared lentivirus to insert the Wnt3a gene. After cultivation in media mixed with 2.5 µL/mL puromycin for an additional 48 hours, the infected cells were identified by GFP expression through confocal microscopy (Carl Zeiss Meditec, Jena, Germany). Wnt3a gene copy number in the MSCs was measured using the Lenti-X Provirus Quantitation kit (Takara Bio).
Experimental animals
Fifty female Sprague-Dawley rats (Orient Bio Inc., Seongnam, Korea), weighing 230-250 g at the beginning of the experiment, were housed in a room with a 12 h/12 h light/dark cycle and free access to food and water.
SCI and experimental groups
Surgical procedures were performed under general anesthesia induced by 1.5% isoflurane (Piramal Critical Care, Inc., Bethlehem, PA, USA). After skin incision, the vertebral column was exposed between T9 and T10, and a laminectomy was performed at the T9 level. The rats received a moderate SCI induced by an Infinite Horizon (IH) impactor (250-kKdyn; Precision System Instrumentation, Fairfax, VA, USA) [27]. After SCI, the contusion site was washed with normal saline, and the skin was sutured. Cefazolin (10 mg/kg) and ketoprofen (5 mg/kg) were subcutaneously administered for antibiotic prophylaxis once a day for 3 days, and each rat was housed individually. The rat’s bladders were emptied manually thrice daily until spontaneous voiding was recovered. At the beginning of the experiment, there were 50 animals, but 14 animals were excluded due to death or injury failure. Rats were blindly separated into five groups : 1) phosphate-buffered saline (PBS) injection only group (injury only group, n=7); 2) hMSC transplantation group (MSC group, n=7); 3) hMSC transfected with pLenti vector (without Wnt3a gene) transplantation group (pLenti-MSC group, n=7); 4) hMSC transfected with Wnt3a gene transplantation group (Wnt3a-MSC group, n=7); and 5) hMSC transfected with enhanced Wnt3a gene transplantation group (1.7 Wnt3a-MSC group, n=8).
Cell transplantation
Six weeks after SCI, all rats were anesthetized with 2% isoflurane. After spinal cord re-exposure, hMSCs were delivered at a rate of 1 µL/min using a 31 G Hamilton microsyringe and an automated microsyringe pump (Harvard Apparatus, Holliston, MA, USA), which are fixed to the stereotactic device. The cells were inserted into the center of the contusion site at a depth of 1 mm from the dorsal dura. The number of injected cells in each group for transplantation was 5×105 cells/15 µL/point at 2 points (the injury epicenter and 2 mm proximal to the epicenter). Total cell numbers were 1×106 cells/30 µL. After injection, the needle was kept in place for 5 minutes to prevent the solution from flowing backward and was then retracted over a subsequent 5 minutes.
Behavioral test
After preparing the rat SCI models, we assessed the neurological function of each rat using the Basso-Beattie-Bresnahan (BBB) locomotor rating scale [1]. Behavioral tests were performed on the first, second, and third day after injury, and then weekly for 14 weeks. Before the BBB test, each rat was allowed to adapt to the open field (90×90 cm, 10 cm wall height). Once the rat demonstrated acclimatization by walking continuously in the open field for 10 minutes, an examiner blinded to group-identifying information assessed the locomotor performance of each rat using the BBB scale. The behavioral test scores of all groups were analyzed by one-way analysis of variance (ANOVA), followed by a post hoc Tukey-Kramer multiple comparison test. All statistical analyses were performed using Statistical Package for Social Sciences (SPSS) ver. 12.0.1 (SPSS Inc., Chicago, IL, USA). Statistical significance was set at p<0.05.
Real-time quantitative polymerase chain reaction
Total RNA was isolated from MSCs using TRIzol Reagent (Thermo Fisher Scientific) in accordance with the manufacturer’s protocol. Total RNA aliquots (2 µg) were then reverse transcribed using a TOPscript cDNA Synthesis Kit (Enzynomics, Daejeon, Korea). Real-time quantitative polymerase chain reaction (qPCR) was performed on a CFX Connect Real-Time PCR Detection System (Bio-rad, Hercules, CA, USA). Gene-specific primer sets were designed, and the following primers were used to amplify the target mRNA : Wnt3a (F) 5′-CATGAACCGCCACAACAAC-3′ and (R) 5′-TCGCAGAAGTTGGGCGA′-3′, β-actin (F) 5′-CTTCGGGAAGGAGCTCAAA-3′ and (R) 5′-TGCCTCGTTGTTGTGCAAG-3′. Gene expression was normalized to that of β-actin, which was used as an internal control. Relative quantities of the mRNA of interest were calculated using the comparative threshold cycles (2−ΔΔCt) method.
Western blotting
Cells were lysed in radioimmunoprecipitation assay (RIPA) lysis buffer (Thermo Fisher Scientific) containing the Halt protease inhibitor cocktail (Thermo Fisher Scientific). Equal amounts of protein (10 µg) were resolved using sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto a nitrocellulose membrane. Each membrane was blocked in 0.1% Tris-buffered saline with Tween-20 (TBST) containing 5% bovine serum albumin (BSA) and incubated with indicated primary antibodies against Wnt3a (Novus Biologicals, Centennial, CO, USA), β-catenin (Cell Signaling Technology, Danvers, MA, USA), and β-actin (Sigma-Aldrich, St. Louis, MO, USA). The blots were developed using Advansta Western Bright ECL HRP Substrate Kits (Advansta, San Jose, CA, USA) and detected using a C-DiGit Blot Scanner (LI-COR, Lincoln, NE, USA).
Immunofluorescence staining
For immunofluorescence analysis, all rats were sacrificed and perfused with PBS and 4% paraformaldehyde (PFA; Sigma-Aldrich) 7 weeks after transplantation. The lesion sites of the spinal cords were extracted and fixed with 4% PFA for 6 hours, followed by incubation with 30% sucrose in PBS overnight. Serial longitudinal spinal cord sections were collected (10 µm thickness) utilizing a Cryotome (Microm, Waldorf, Germany) and mounted on poly-L-lysine-coated Superfrost Plus slides (Matsunami, Osaka, Japan). The sections were washed three times with Tris-buffered saline (TBS), then blocked with TBS containing 5% BSA and 0.1% Triton X-100 for 1 hour at room temperature. The sections were incubated overnight with primary antibody at 4°C and washed three times with TBS, then incubated with fluorescent-conjugated secondary antibody (1 : 1000; Invitrogen, Carlsbad, CA, USA). The primary antibodies used were : neurofilament (NF, 1 : 200; Abcam, Cambridge, UK), growth-associated protein 43 (GAP43, 1 : 200; Abcam), and microtubule-associated protein 2 (MAP2, 1 : 200; Abcam). Fluorescent images were obtained using a confocal microscope (Carl Zeiss, Oberkochen, Germany) with ZEN microscope software (Carl Zeiss). Fluorescent intensity was analyzed using ImageJ program (National Institutes of Health and the Laboratory for Optical and Computational Instrumentation (Madison, University of Wisconsin).
Cytokine analysis
After 48 hours incubation in opti-MEM medium (Gibco; Thermo Fisher Scientific), supernatants of MSC, hMSC transfected with Wnt3a gene, hMSC transfected with enhanced Wnt3a gene were collected. Supernatants were biotin-labelled and hybridized onto human L1000 glass slide arrays (RayBiotech, Peachtree Corners, GA, USA). Detected signals were then normalized using BCA protein assay kit.
RESULTS
Development of enhanced Wnt3a-MSC
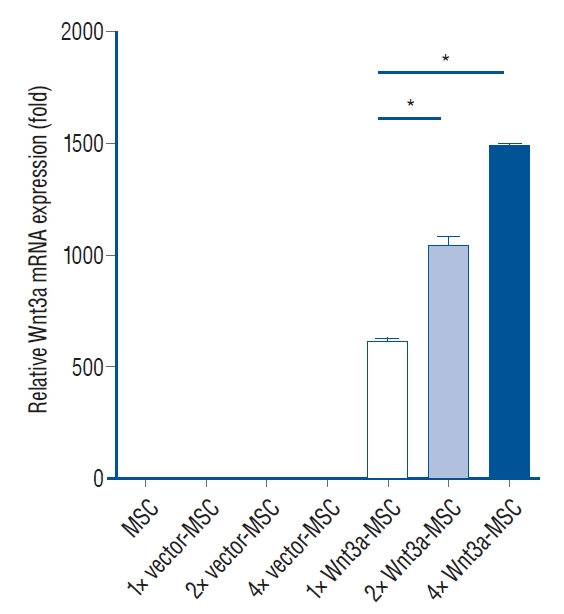
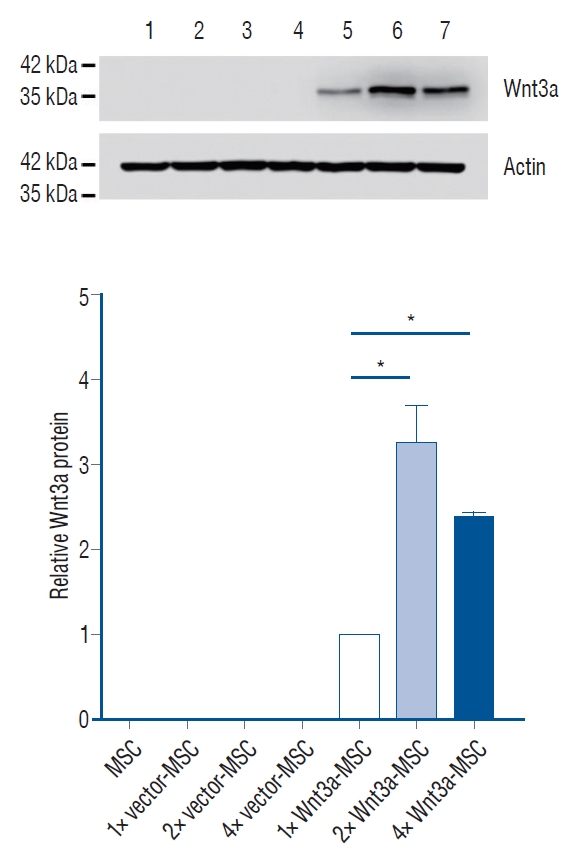
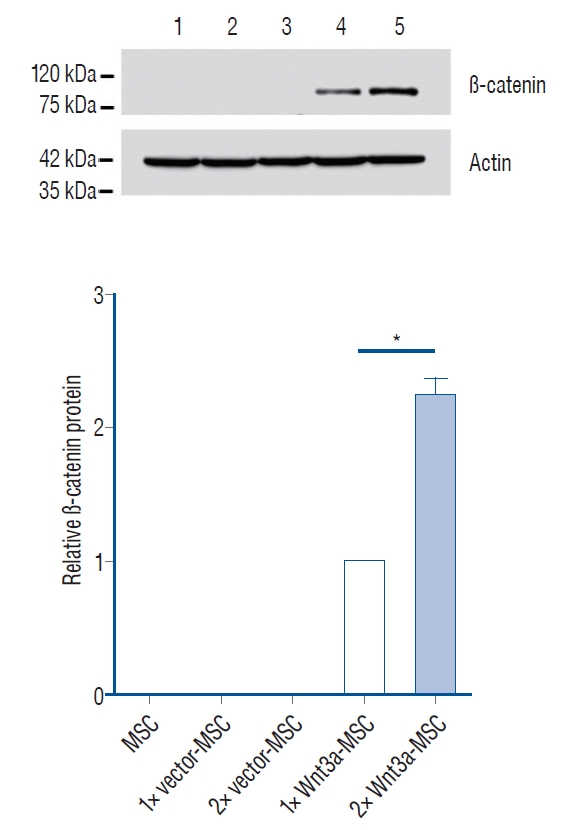
In a previous study, we manufactured Wnt3a-expressing MSC (baseline gene expression per cell), but these cells could not statistically improve behavioral results when compared to pure MSC (data not shown). Thus, we needed an improved Wnt3a-MSC that could secret more Wnt3a protein, allowing for more effective signaling for axon regeneration. We developed a new Wnt3a-MSC by increasing the quantity of Wnt3a gene for transfecting into MSC (2 fold and 4 fold). The results shown in Fig. 1 indicate that 2 fold Wnt3a gene transfected MSC (2× Wnt3a-MSC) showed 1.7 fold Wnt3a mRNA expression level as compared to baseline Wnt3a gene transfected MSC (1× Wnt3a-MSC), while 4 fold Wnt3a gene transfected MSC (4× Wnt3a-MSC) showed 2.4 fold Wnt3a mRNA expression level as compared to 1× Wnt3a-MSC. Interestingly, we found that 2× Wnt3a-MSC secreted more Wnt3a proteins even compared to the 4× Wnt3a-MSC (Fig. 2); the 2× Wnt3aMSC showed 3.25 fold Wnt3a protein level as compared to 1× Wnt3a-MSC (Fig. 2). The 2× Wnt3a-MSC also showed a 2.2 fold increase in β-catenin expression level when compared to 1× Wnt3a-MSC (Fig. 3). Given these observations, we selected 2× Wnt3a-MSC to be the new enhanced Wnt3a-MSC, henceforth referred to as 1.7 Wnt3a-MSC. In the 1× Wnt3a-MSC, 3.5×104 copies of the virus were utilized per MSC, whereas in the 1.7 Wnt3a-MSC, 7×104 copies of virus were utilized per MSC. After each round of transfection, we quantified the provirus copy number using the Lenti-X Provirus Quantitation kit. We detected 13.02 provirus copies/cell in 1× Wnt3a-MSC, and 22.19 provirus copies/cell in 1.7 Wnt3a-MSC.
Enhanced Wnt3a gene expression in hMSC promotes behavioral recovery
We injected PBS (injury only group), hMSC (MSC group), hMSC transfected with pLenti vector (pLenti-MSC group), hMSC transfected with Wnt3a genes (Wnt3a-MSC group), and hMSC transfected with enhanced Wnt3a genes (1.7 Wnt3a-MSC group) in the respective groups 6 weeks after SCI. We found that the 1.7 Wnt3a-MSC group showed significant behavioral recovery compared with all other groups (p<0.05). BBB score of the 1.7 Wnt3a-MSC group (15.0±0.28) was significantly higher than that of the injury only (10.0±0.48), MSC (12.57±0.48), pLenti-MSC (12.42±0.48), and Wnt3a-MSC (13.71±0.61) groups (Fig. 4).
Immunofluorescence for axonal regeneration markers
The expression of axonal regeneration markers was analyzed in a region proximal to the injury epicenter, where cells were injected. Confocal imaging revealed that the expression of axonal regeneration markers such as GAP43, MAP2, and NF were increased in the Wnt3a-MSC and 1.7 Wnt3a-MSC groups as compared with injury only, MSC, and pLenti-MSC groups (Figs. 5-7). However, it was difficult to find a significant difference between the Wnt3a-MSC and 1.7 Wnt3a-MSC groups (Figs. 5c, 6c, and 7c).
DISCUSSION
We previously demonstrated the therapeutic effects of MSC treatment on neural regeneration via direct injection of cells into injured spinal cords of chronic SCI patients [16,20]. However, in these two clinical trials, pure MSC did not show sufficient effects even though it demonstrated safety in the long-term follow-ups. Therefore, to maximize the neural regenerative effect through cell transplantation, we developed enhanced MSCs that secrete Wnt3a protein. There was also a need to calibrate the accuracy of the chronic SCI model in animals. We tested the efficacy of Wnt3a-secreting cell transplantations in regenerating injured axons in a rat model of SCI, and found a significant degree of axonal regeneration and recovery of motor function [21,25,26]. However, there was a discrepancy between the clinical and animal studies. In the clinical studies, MSC were injected in chronic SCI patients. In the animal studies, we could not be certain whether the animal model used in the previous study was at the chronic or subacute stage of SCI. Although the time point used, i.e., 4 weeks after SCI, is generally regarded as chronic SCI as per previous animal studies [15], we could observe that slight gradual BBB increase during 2 weeks even after 4 weeks of SCI. On the contrary, the clinical study performed with the definition of the chronic phase as after 12 months of injury with no neurological changes in the previous 6 months [6,9]. Therefore, in this study, we decided upon the time point of 6 weeks after SCI in a rat model of chronic SCI, based on our previous pilot study (data not shown).
Several mechanisms behind the effectiveness of stem cell transplantation in recovering SCI have been proposed, such as differentiation of spinal cord neurons leading to tissue preservation, decreased injury size, and increased axonal regeneration [11,17,19,24]. Secreted trophic factors from MSC, such as nerve growth factor, vascular endothelial growth factor, and brainderived neurotrophic factor, are known to promote axonal regeneration and functional recovery as well [4,24]. It is also possible that genetically engineered neural stem cells can exhibit better spinal cord repair and cell survival, and migrate longer distances [2,5,11]. In the present study, we modified MSC with the Wnt3a gene to enhance the function of pure MSC. Whereas endogenous Wnt1 and Wnt5a signaling mediates only limited functional recovery after SCI [7,13], canonical Wnt3a signaling promotes spinal cord neural precursor development and stimulates axonal regeneration [22,26]. We have also demonstrated that Wnt3a by itself, Wnt3a-secreting fibroblasts, and Wnt3a-secreting fibroblasts with alginate scaffolds can improve functional recovery in SCI rat models [21,26]. In this study, stem cell injection was performed using a stereotactic device and a microsyringe pump (1 µL/min), and only a Hamilton syringe (31G) was inserted into the tissue, and there were no side effects. In most of the animal studies, the injection method using a stereotactic device and a microsyringe pump is used, and in clinical studies, manual injection was also performed to the patient [10,20]. In addition, regarding the rationales of the 2 point injection, we reported at the previous paper : the cavity usually developed in the injury epicenter, which has low tissue pressure. Therefore, when we inject the MSC in the injury epicenter, the cells are kept in the tissue due to low tissue pressure, but there is a disadvantage of low cell survival from lack of blood circulation. On the contrary, to inject cells at 2 mm proximal to epicenter has an advantage in cell survival, but there is a possibility of cell leakage [20].
We hypothesized that increasing the level of gene expression of Wnt3a in hMSC could highly promote axonal regeneration in injured spinal cords as compared to low-expressing Wnt3atransfected hMSC. Hence, we developed a new Wnt3a-MSC which was transfected with the Wnt3a gene (2×, 4×). We selected the 2× Wnt3a-MSC as an enhanced Wnt3a-MSC, and referred to it as 1.7 Wnt3a-MSC because it showed 1.7 fold Wnt3a mRNA expression. The 1.7 Wnt3a-MSC showed higher β-catenin expression level when compared to baseline (1× Wnt3a-MSC). A result of western blot of Wnt3a protein showed that 2× Wnt3a-MSC expressed more Wnt3a protein than 4× Wnt3a-MSC (Fig. 2). These data suggest that protein expression does not increase proportionally when mRNA is increased, and that the optimal volume of mRNA in the cell exists. It is known that the amount of mRNA and that of produced protein are not always proportional to each other, and the underlying mechanism in translation seems to be affected to the concentration of ribosomes, the effect of tRNA, and protein characteristics. In this study, we found out the optimal rate of Wnt3a mRNA is 1.7 Wnt3a-MSC (2× Wnt3a-MSC) [12]. We finally found that the 1.7 Wnt3a-MSC showed the best titration result showing the best BBB improvement. In addition to Wnt3a, the cells of MSC, Wnt3a-MSC and 1.7 Wnt3a-MSC groups released different profiles of other neurotrophic factors (Table 1). The cells of Wnt3a group produced more secretions than the cell of MSC group in brain-derived neurotrophic factor (BDNF), epidermal growth factor (EGF), fibroblast growth factors (FGF)-4, FGF-7, glial cell line-derived neurotrophic factor (GDNF), neurotrophin (NT)-4, and the cells of 1.7 Wnt3a group secreted more than the cells of Wnt3a group in BDNF, Beta-FGF, Beta-NGF, FGF-7, NT-3, NT-4, indicating that the three cytokines (BDNF, FGF-7, NT-4) especially showed linear increase in MSC, Wnt3a, and 1.7 Wnt3a groups, and they seemed to be persistently related to spinal cord regeneration (Table 1). It is well-known that BDNF, GDNF, NGF, and NT-3 are mainly effective neutrophic factors for spinal cord regeneration [3]. This information proved valuable when we applied the enhanced Wnt3a-MSC to SCI animal model. Therefore, we could name the 1.7 Wnt3a-MSC as a “smart stem cell” regarding functional recovery in SCI. However, since it is the result of one media analysis, statistical processing was not conducted, and statistical verification should be performed through multiple media analyses in the further study.
We performed BBB test to demonstrate functional recovery from SCI after injection of hMSC with enhanced Wnt3a gene expression. The 1.7 Wnt3a-MSC group showed significantly more improvement than other groups at 8 weeks after transplantation. Fourteen weeks after injury (8 weeks after transplantation), the scores of all groups plateaued. Thus, we selected this time point to be representative of final motor recovery and reasonable to assess locomotor improvement. In this study, we did not evaluate neuropathic pain, such as the von Frey test, but did only motor power recovery. In the further study, we are needed to investigate the neuropathic pain and bladder function recovery that are clinically common problems in chronic SCI. This will be a multiple aspect study for spinal cord regeneration.
Immunofluorescence was performed for visual analysis of axonal regeneration. GAP43, MAP2, and NF are well known representative markers of axonal regeneration. These markers were used to determine whether the increased behavioral test scores were the result of axonal regeneration or other neuroprotective effects. Because both the 1.7 Wnt3a-MSC and the Wnt3a-MSC groups showed higher expression of all axonal regeneration markers than the other groups, we can conclude that the BBB improvement was from axonal regeneration. However, we could not differentiate the differences in the staining of markers between the effects of 1.7 Wnt3a-MSC and Wnt3a-MSC injection in these rats in this study (Figs. 5-7). We speculate that enhanced Wnt3a gene MSC (1.7 Wnt3a-MSC) can exert stronger regeneration effect for injured spinal cord, and the result was evaluated by behavior test (BBB score), but unfortunately immunostaining with GAP43, MAP2, and NF showing spinal cord regeneration failed to demonstrate the precise difference of axon regeneration between Wnt3a-MSC and 1.7 Wnt3a-MSC group. To demonstrate the effect of spinal cord regeneration by Wnt3a gene enhancing with imaging technique, we need to additional imaging method such as axon tracing in further study. Basically, this study was investigated in the chronic phase of SCI which inflammation and tissue necrosis has already disappeared, and glial scar formation was set, in which neurological deficit only exists unlike phenomenon in acute or subacute phase. Therefore, analysis on the effect of angiogenesis, anti-inflammation, and glial scar formation, which are important mechanisms of neuroprotection in acute or subacute phase of SCI, was not done to focus on the recovery mechanism for chronic status of SCI. In addition, we did not conduct the analysis of lesion cavity size or fibrosis change, which are another aspect to figure out the spinal cord regeneration. We consider that this is the limitation of our study for histology, and we are needed to add them in further study.
CONCLUSION
We produced 1.7 fold Wnt3a mRNA expressing hMSCs to promote the efficacy of axonal regeneration based on the safety of pure hMSCs and found that this gene titration is optimal to BBB improvement through axonal regeneration. Our study suggests that enhanced gene expression with optimal ratio in gene transfected MSC can significantly impact axonal regeneration and functional recovery in a rat model of chronic SCI more than pure hMSC.