INTRODUCTION
Spinal cord injury (SCI) can cause clinically irreversible disability and result in much comorbidity. The primary SCI is direct injury from an initial mechanical trauma, and the secondary injury results from progressive processes that augment the injury resulting in a protracted period of tissue destruction1,2,29). These cascading injuries make recovery from SCI hard or irreversible. However, recent studies demonstrated that spontaneous neuronal regeneration can occur in rat models of SCI15,16,19). Existence of neural stem/progenitor cells (NSPCs) in adult stage was proven in adult mammals, including humans11,17). In particular, there were many reports that SCIs induce proliferation and expression of spinal NSPCs. These observations suggest that adult NSPCs may work for neuronal regeneration in adult mammals following SCI.
Valproic acid (VPA) is widely used for the treatment of seizures and bipolar disorders. In addition, VPA is a potent histone deacetylase (HDACs) inhibitor, which is critical to cellular inflammatory and repair processes8). In many animal model studies of neurodegenerative diseases, VPA has beneficial effectsin treatment of stroke, amyotrophic lateral sclerosis, spinal muscular atrophy, Parkinson's disease and Alzheimer's disease7,10,20,27,28,32,33). Recently, VPA was shown to be important for expression and self-renewal of hematopoietic stem cells6). We hypothesized that VPA can stimulate expression of NSPCs. Therefore, this study is intended to investigate the effects of VPA on NSPCs expression in a rat SCI model.
MATERIALS AND METHODS
Animal surgery and administration of VPA
All animal experiments were performed in accordance with the National Institutes of Health guidelines on animal care, and were approved by the Institutional Animal Care Committee. All efforts were made to minimize the number of animals used and animal suffering. Adult male Sprague-Dawley rats (n=30) weighing 290-310 grams (Samtako Bio, Osan, Korea) were randomly and blindly allocated into three groups. In group 1 (sham, n=8), laminectomy was performed. In group 2 (SCI-VPA, n=11), the animals received a dose of 200 mg/kg of VPA (Sigma-Aldrich, St. Louis, MO, USA). In group 3 (SCI-saline, n=11), animals received 1.0 mL of the saline vehicle solution. Rats were anesthetized intraperitoneally with a mixture of xylazine (10 mg/kg) and ketamine (60 mg/kg). After laminectomy at T9, the extradural plane between the dura and adjacent vertebrae was carefully dissected. A modified aneurysm clip with a closing force of 30 grams (Aesculap, Tuttlingen, Germany) was held in an applicator in the open position. The clip was rapidly released from the applicator and applied vertically onto the exposed spinal cord for a 2-minute compression. For the sham controls the same surgical procedure was followed, but clip compression was not applied. After surgery, the muscle, fascia, and skin were sutured using a 4-0 silk suture. Rectal temperature was maintained at 37.0┬▒0.5Ōäā by a thermostatically-regulated heating pad during surgery, and during recovery, animals were placed overnight in a temperature and humidity controlled chamber. To reduce post-surgery isolation-induced stress, rats were housed in pairs at an ambient temperature of 22-25Ōäā in an alternating 12-hour light/dark cycle. Bladders were manually emptied twice daily until spontaneous voiding occurred. At three days after surgery, we checked functional deficit using the open locomotor rating scale by Basso, Beattie, and Bresnahan (the BBB score)3). All rats showed 4 or 5 BBB score, indicating proper cord damage in SCI model. A dose of 200 mg/kg of VPA or normal saline as a vehicle control was intraperitoneally injected twice daily at 12 hours intervals for 7 days. The total daily VPA dose of 400 mg/kg/day was similar to doses used in previous studies9,34). To evaluate histological changes, the animals were sacrificed and the spinal cords were collected 1 week after SCI. For immunohistochemistry analysis, samples were prepared from the sham group (n=5), SCI-saline group (n=7), and SCI-VPA group (n=7). Samples for western blotting analysis were prepared from rats in the three groups (group 1=3 rats, group 2, 3=4 rats).
Immunohistochemistry of nestin
Rats were deeply anesthetized by an intraperitoneal injection of ketamine and were perfused intracardially with 4% paraformaldehyde in 0.1 M sodium phosphate buffer (PB, pH=7.4). The thoracic spinal cord was excised, postfixed for 24 hours, and maintained overnight in 30% sucrose in 0.1 M PB. Spinal cord tissues were sectioned at a thickness of 30 ┬Ąm on a cryostat, and sections were floated on the surface of 0.1 M PB. A 5 and 6 mm section rostral to the center of injury was selected. To detect nestin (marker for neural stem cell), spinal cord sections were blocked with 4% normal serum in 0.5% Triton X-100 for 1 hour at room temperature and incubated overnight at 4Ōäā with a 1 : 2000 dilution of mouse monoclonal anti-nestin (R&D Systems Inc., Minneapolis, MN, USA), and rinsed for 3├Ś10 min in 0.1 M PB. Sections were then incubated in 0.1 M PB containing 4% normal serum and 0.5% Triton X-100 for 2 hours at 25Ōäā on a shaker, and then in primary antiserum in 0.1 M PB containing 4% normal serum and 0.5% Triton X-100 for 12 hours at 25Ōäā. After rinsing (3├Ś10 min) in 0.1 M PB, sections were incubated in a 1 : 200 dilution of biotinylated anti-mouse IgG (Sigma, St. Louis, MO, USA) in 0.1 M PB containing 4% normal serum and 0.5% Triton X-100 at 25Ōäā for 2 hours. The sections were then incubated in a 1 : 50 dilution of avidin-biotinylated horseradish peroxidase (Vector Laboratory) in 0.1 M PB for 2 hours and rinsed (3├Ś10 min) in 0.25 M Tris. Finally, staining was visualized by reaction with 3, 3'-diaminobenzidine tetrahydrochloride (DAB) and hydrogen peroxide in 0.25 M Tris for 3-10 min using a DAB reagent set (Kierkegaard&Perry, Gaithersburg, MD, USA). All sections were rinsed in 0.1 M PB and mounted on Superfrost Plus slides (Fisher, Pittsburgh, PA, USA) and dried overnight at 37Ōäā. The mounted sections were dehydrated with alcohol, cleared with xylene, and cover slipped with Permount mounting medium (Fisher). The labeled cells were identified and counted with separation of antibody at three tissues in each different animal. The labeled tissues were photographed using a Zeiss Axiopan microscope with high power DIC optics (Carl Zeiss Meditec Incorporation, Jena, Germany). The images were viewed on a computer monitor using a Zeiss Plan-Apochromat 40x objective (Carl Zeiss) and photographs of the central canal region and ventral side of white matter of left and right sides were taken with a Zeiss AxioCam HRc digital camera (Carl Zeiss). Enumeration of immune-positive cells used a Labworks, version 4.5, computer-assisted image analyzer (UVP, Upland, CA, USA).
Western blotting of SOX2
Rats in three groups were decapitated rapidly under anesthesia. The thoracic spinal cord was rapidly dissected and then immediately frozen in liquid nitrogen. Frozen tissue was mixed with RIPA buffer (25 mM Tris-Cl pH 7.6, 150 mM NaCl, 1% NP-40, 1% sodium deoxycholate, 0.1% SDS) containing protease inhibitor cocktail (Roche, Mannheim, Germany) and immediately homogenized. The homogenate was centrifuged at 13000 rpm for 30 min at 4Ōäā and the supernatants were determined using the BCA protein assay (Sigma, St. Louis, MO, USA). Proteins were separated by 10% SDS-PAGE gel and transferred to a nitrocellulose membrane. After incubation in a blocking solution of 5% non-fat dry milk in Tris-buffered saline containing 0.1% Tween-20 for 1 h at room temperature. The membrane was incubated with a 1 : 1000 dilution of mouse monoclonal anti-SOX2 (cell signaling) and mouse monoclonal anti-╬▓-actin (Sigma-Aldrich), and overnight at 4Ōäā, and then with a horseradish peroxidase (HRP)-conjugated secondary antibody for 1 h at room temperature. The proteins were detected with chemiluminescence reagents. Immune-positive bands used an image J, version 1.46r, computer-assisted image analyzer (National Institutes of Health, USA).
Statistical analysis
All statistical comparisons were computed using SPSS 17.0 (SPSS, Inc., an IBM Company, Chicago, IL, USA). Data are expressed as mean┬▒standard error of the mean. Repeated measure ANOVA was used to compare groups. Null hypotheses of no difference were rejected if p-values were less than 0.05.
RESULTS
Nestin expression in SCI
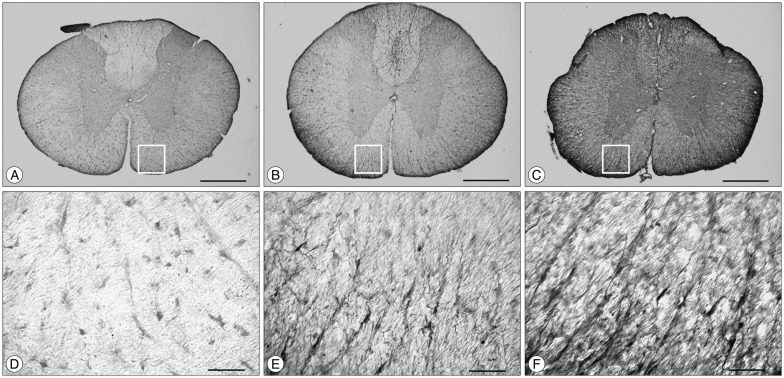
We analyzed the nestin immunoreactivity from two different sites of the spinal cord based on methods from Sibuya et al.32). One site is the white matter of ventral side and the other is the ependyma of central canal in a cross-section of spinal cord. We used ANOVA analysis for confirmation of density areas of nestin.
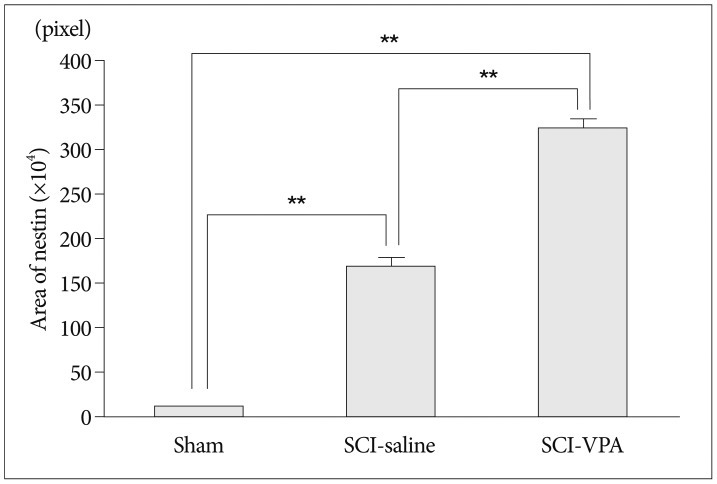
In the white matter of ventral side, nestin immunoreactivity was almost undetectable in the sham group (Fig. 1A, D). SCI groups showed remarkable nestin immunoreactivity compared with sham group. In the SCI-saline group, nestin immunoreactivity extended in arboroid processes from the pial surface toward the spinal cord center (Fig. 1B, E). In the SCI-VPA group, nestin immunoreactivity was much stronger than in SCI-saline group (Fig. 1C, F). Density areas of nestin expression in the white matter of ventral side differed significantly between the SCI and sham group (Fig. 2). The SCI group showed significantly large density area compared with sham group and within SCI groups, the SCI-VPA group had a much larger density area of nestin than the SCI-saline group.
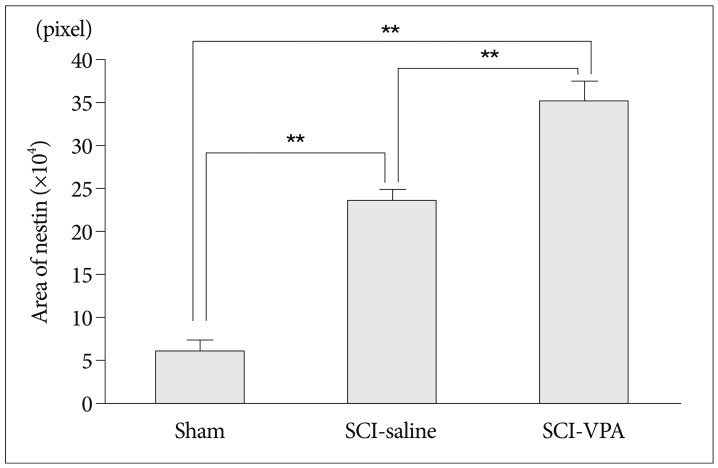
In the ependyma of the central canal, nestin immunoreactivity also was strongest in the SCI-VPA group (Fig. 3C). In sham group, nestin immunoreactivity was observed in some cells (Fig. 3A). SCI groups had significant nestin immunoreactivity compared to the sham group and extended in processes from the whole ependymal area (Fig. 3B, C). The density areas of nestin expression in the ependyma of the central canal were similar to the white matter of ventral side. The SCI groups had significantly large density areas of nestin compared to the sham group, and SCI-VPA group had a much larger density area of nestin than SCI-saline group (Fig. 4).
To sum up, VPA increased expression of nestin, a marker for NSPCs, in the white matter of the ventral side and ependyma of the central canal.
SOX2 analysis in SCI
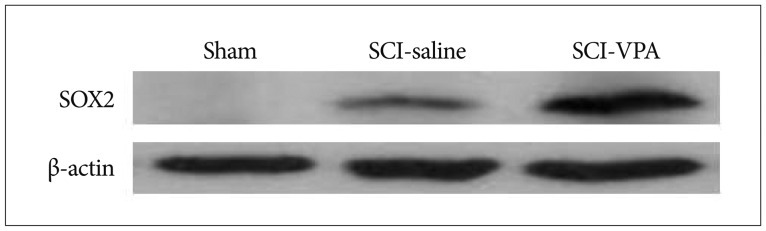
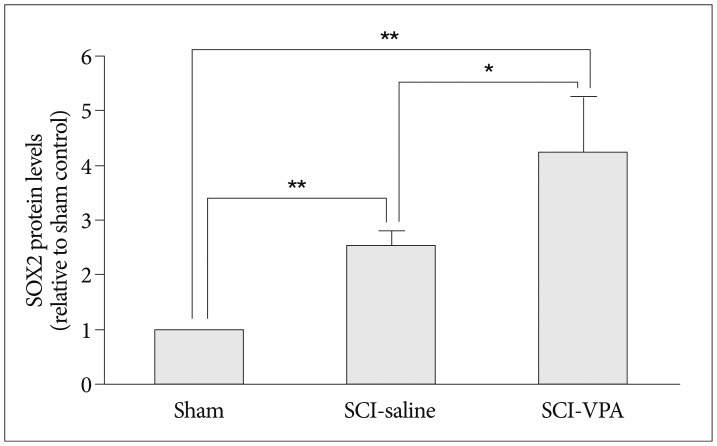
SOX2 was detectable in SCI groups and undetectable in sham group. Besides, in SCI groups, VPA significantly increased the SOX2 positive band than in the SCI-saline group (Fig. 5). This difference was confirmed by ANOVA analysis. SOX2 protein levels were highest in SCI-VPA group, followed by SCI-saline (Fig. 6).
DISCUSSION
Recovery from SCI is a major goal of neurosurgeons. Early decompressive surgery or high-dose steroid therapy, while long-standing treatments of choice, do not provide recovery from critical sequelae after SCI. Efforts to minimize secondary injury of SCI have included drug management as a neuroprotective effect in many recent studies, including minocycline, erythropoietin, and Nogo-66 receptor antagonist13,23,24). VPA also was investigated35). These drugs are efficacious for minimizing scarring and cavitation caused by SCI13,23,24,35). However, the so-called "neuroprotective effect" was not enough for radical management of SCI. The ultimate goal of management for SCI is regeneration of injured neural tissues, so numerous studies of neural stem cell therapy have been reported.
Embryonic stem cells are pluripotent and self-replicating. Transplantation of human embryonic stem cells to injured rat spinal cord could result in recovery26). Despite several limitations to be overcome, including differentiation to purified neural cell type5,18,26) and teratoma formation4), human embryonic stem cell therapy is an attractive method for recovery from SCI. This strategy has been recently validated21). The existence of NSPCs at the embryonic stage and at the adult stage is already proven11,17). Reported adult NSPCs so far are the ependymal cells and subependymal cells of the cerebral ventricles and glial fibrillary acidic protein-positive cells in the subventricular zone. Interestingly, SCI induces expression of NSPCs and this phenomenon may be associated with neuronal repair and regeneration after SCI30).
Various markers for NSPCs are reported, and among these we used nestin and SOX2. Nestin, an intermediate filament protein, is a widely employed marker of multipotent neural stem cells in adult CNS. In an experiment on an SCI rat model, nestin expression increased time-dependently, and peaked in the 5-mm rostral region to the injury center at 1 week after SCI30). SOX2, sex-determining region Y-box 2, is a transcription factor that is essential for regulation of self-renewal and potency of embryonic and adult neural stem cells and express in the adult multipotent NSPCs22). In an experiment using SOX2 as NSPC's marker on SCI rat model, it is observed that SOX2 expression reaches a peak at 1-2 weeks after SCI22). Based on this prior research, we selected the killing time and the region of transection, at one week after injury and 5-mm rostral to injury.
VPA, 2-propylpentanoic acid, is an HDAC inhibitors. HDAC enzymes can be classified into four major classes according to their homology to yeast HDACs8). VPA inhibits HDAC8, a class 1 HDAC8). HDAC inhibitors are involved in normalization of histone hypoacetylation and transcriptional dysfunction of various neurodegenerative conditions8). The neuroprotective effects of treatment with VPA as HDAC inhibitor are proven in various animal models of neurodegenerative diseases, such as stroke, amyotrophic lateral sclerosis, spinal muscular atrophy, Parkinson's disease and Alzheimer's disease7,10,20,27,28,32,33). Furthermore, we reported that VPA minimizes secondary injury of SCI by diminishing cavitation volume inflammatory reactions, and restoring the hisone acetylations in injured spinal cords after SCI in our previous study35). A recent study demonstrated that VPA as HDAC inhibitor has the ability to stimulate proliferation and self-renewal of hematopoietic stem cells (HSCs)6). In above study, it was reported that VPA increases the proliferation of human CD 34+ HSCs but does not induce differentiation. In addition, VPA activates glycogen synthase kinase 3╬▓ (GSK3╬▓)-dependent signaling pathways necessary for the self-renewal potential of HSC. VPA induces proliferation of neural progenitor cell by activation of Wnt/GSK3╬▓/╬▓-catenin pathway in rat brain12). These findings prompted to us to investigate the effect of VPA on expression of NSPCs in SCI rat model. In our study, VPA increases the expression of NSPCs after SCI. This result is similar to previous studies6,12), but, in another study, activation of GSK3╬▓ by phosphorylation promotes differentiation of NSPCs and reduces proliferation of NSPCs in rat subventricular zone25). Weighing this evidence, it is certain that the GSK3╬▓ signaling pathway is important for proliferation of NSPCs. Therefore, we guess that the effect of activated GSK3╬▓ signaling pathway by VPA may be different according to circumstance. There have been no studies so far about VPA effect on NSPCs expression in SCI model as in our study. Further studies are needed to investigate how VPA causes expression of NSPCs by GSK3╬▓ signaling pathway.
Furthermore, there is one thing to be concerned about. As known by previous studies, VPA contributes to an anti-inflammatory reaction by down-regulation of various inflammatory genes31) and this anti-inflammatory reaction is conducive to migration of adult NSPCs to the injury site14). Consequently, we can draw a hypothesis that the above sequential steps cause VPA to negatively affect the reaction for migration of NSPCs to the injury site. However, in our study, we cannot tell how much VPA affects migration of NSPCs to the injury site. According to our results, it is certain that VPA can increase expression of NSPCs around the injury site. Because existence of endogenous NSPCs were already proved in the spinal cord by a previous study30), we would suggest one hypothesis : a higher proportion of assignment for NSPCs expression may at increased proliferation of endogenous NSPCs more than at decreased migration of NSPCs by VPA in the case of SCI.
There is no previous in vivo study about the VPA effect on migration of NSPCs to the injury site. In a future study, we have to launch an in vivo SCI model study using VPA for investigation of the effect on migration of NSPCs.
CONCLUSION
In our study, nestin and SOX2 as markers for NSPCs showed increased expression in the SCI-VPA group in comparison with the SCI-saline group. This result indicates that VPA increases expression of spinal NSPCs in SCI. In future research, we should evaluate the GSK3╬▓ signaling pathway in spinal cord and phenomenon between proliferation and differentiation of NSPCs in SCI and launch an in vivo SCI model study using VPA for investigation of the effect on migration of NSPCs.